


Visit Human Optimization We are transferring the store over to our new brand called Human Optimization

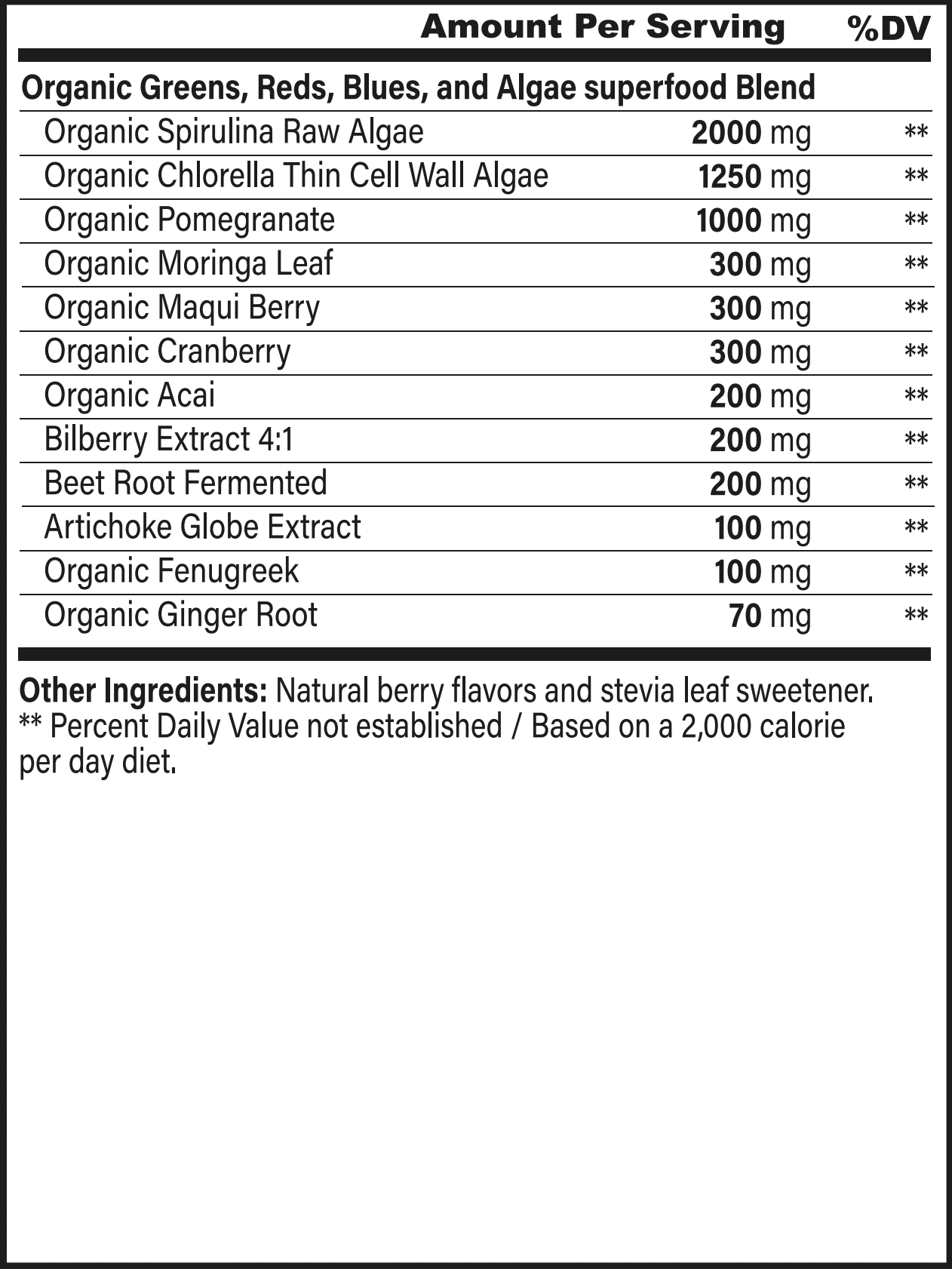
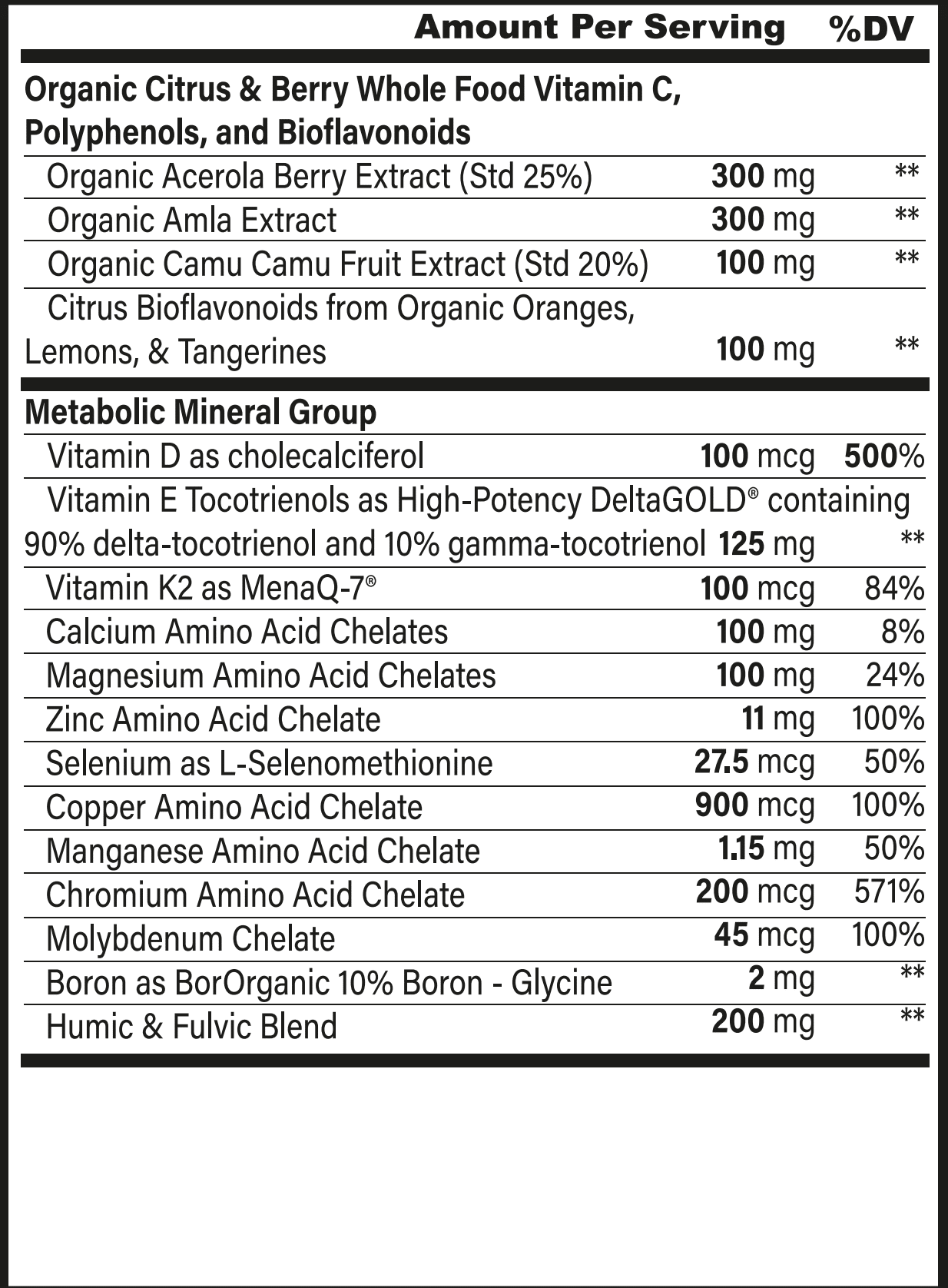
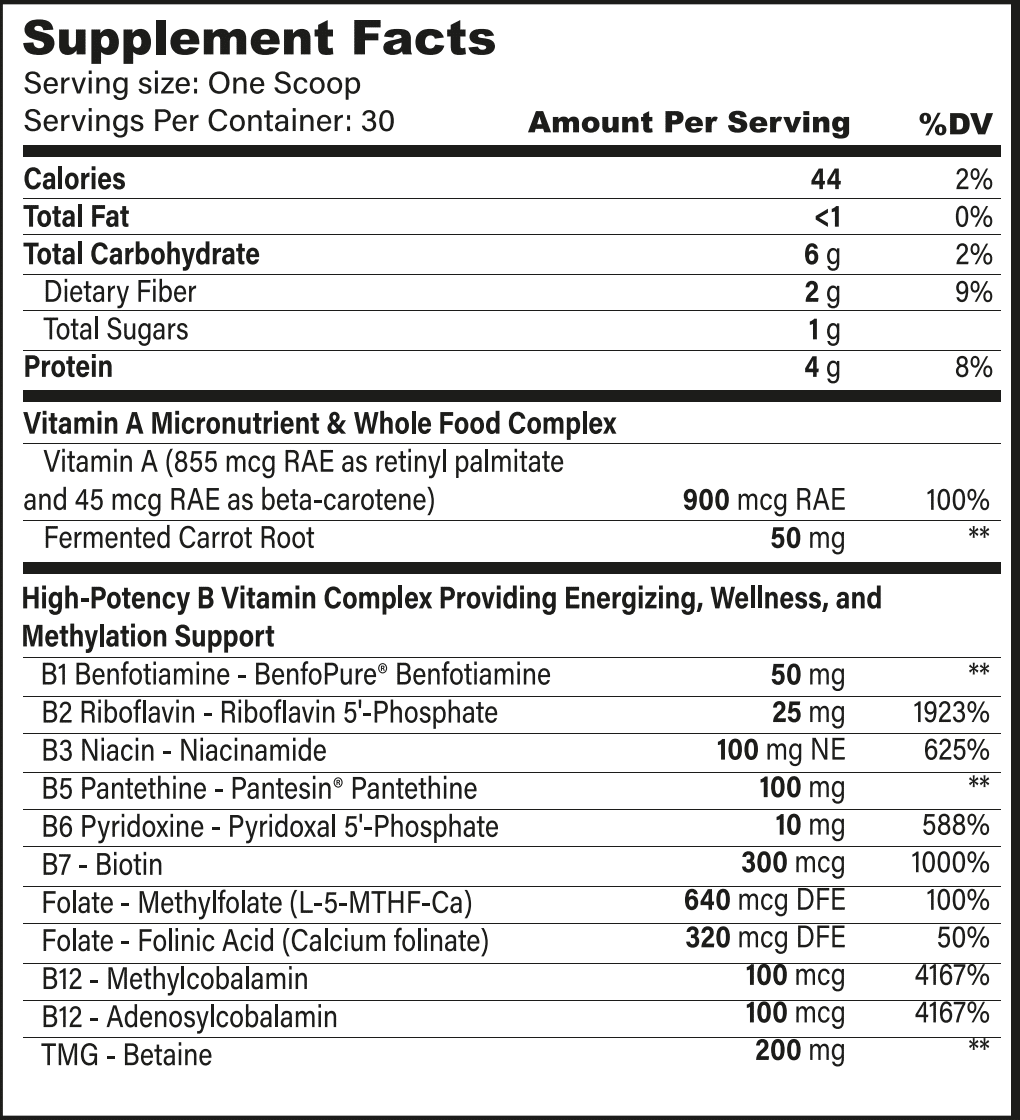
Superfoods, Supplements
Energy Essentials and Superfoods: Subscribe & Save
Original price was: $100.00.$75.00Current price is: $75.00. / month
Related products
-

-
 Energy Support
Energy SupportEnergenesis: 3 Month Supply
Original price was: $357.00.$299.00Current price is: $299.00. Read more -

-
 Brain Support
Brain SupportUltraBrain: Subscribe & Save
Original price was: $99.00.$74.00Current price is: $74.00. / month Read more