


The most advanced and comprehensive multivitamin/multimineral/superfoods blend on the market.


The most advanced and comprehensive multivitamin/multimineral/superfoods blend on the market.
Are your fatigue issues caused by nutritional deficiencies? It's likely at least a partial contributor!
Did you know that your body REQUIRES over 30 different vitamins and minerals and that almost EVERYONE is deficient in at least a few of those compounds?
So what do you need to do?
Simple: Plug the holes in your diet and provide your body with the nutrients and cofactors it needs for optimal health, disease prevention, and energy production. That is exactly what Energy Essentials & Superfoods is designed to do. This is your way to eliminate your deficiencies and virtually guarantee your body has everything it needs to function optimally — every single day, in just 30 seconds.
Introducing Energy Essentials & Superfoods
With 45 bioavailable ingredients and 36 scientifically proven health benefits, it’s the most advanced and comprehensive multivitamin/multimineral/superfood blend on the market
Back to the problem...
In the U.S., 9 out of 10 adults do not meet the recommended daily allowances (RDAs) for Vitamin D and Vitamin E, and over 50% of people don’t meet the RDAs for vitamin A, calcium and magnesium. And that’s just to name a few of many important compounds that many people are deficient in!
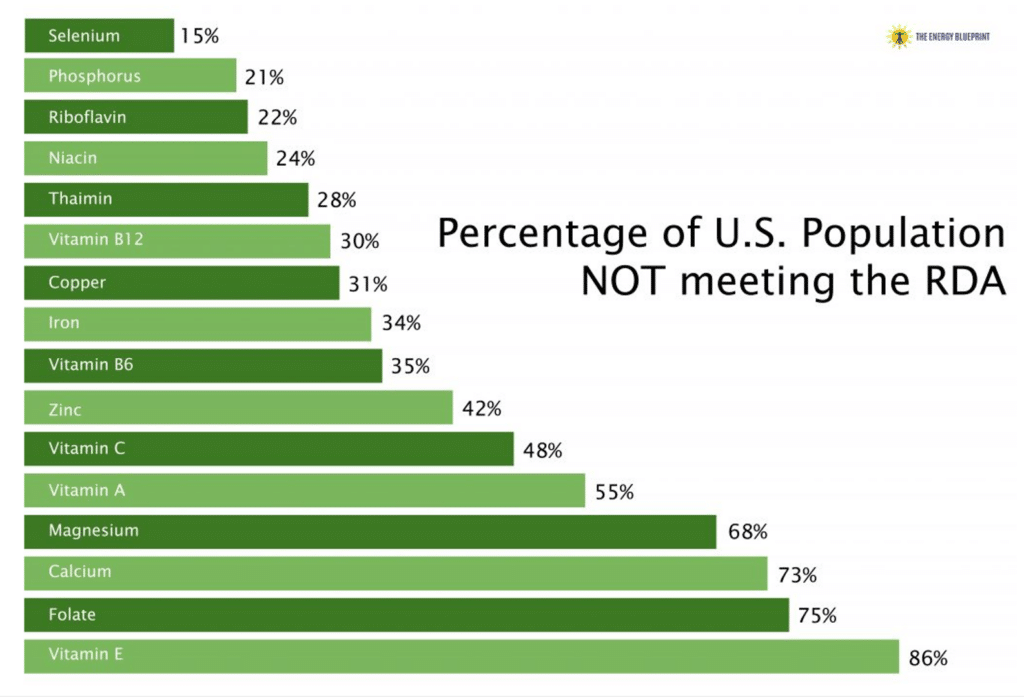
As you can see from this graph, it’s almost guaranteed that you have at least a couple nutrient deficiencies, if not numerous of them. According to the USDA, over 92% of people have deficiencies in essential micronutrients needed to maintain basic health. This is the single most widespread risk factor for chronic disease.
There are 3 major reasons why people aren't getting enough...
- Non-optimal food choices. The most common and most obvious cause of deficiencies is that the majority of people are making very poor food choices — eating a highly processed, unhealthy diet that’s heavy in fats, sugars, and chemicals and light in essential nutrients.
- Poor gut health, stress, alcohol, medications and health issues. Due to poor gut health and microbiome imbalances, many people have absorption problems. So, they can’t even extract the nutrients from their food properly. Plus, many people have chronic stress, chronic infections and chronic medical conditions that rapidly deplete levels of critical vitamins and minerals. Stress alone has been known to deplete vitamins B1, B2, B3, B5, B6, B7, B9, B12, vitamin A, vitamin C, vitamin D, magnesium, chromium, copper, iodine, potassium, selenium, zinc, and more.
- Lower vitamin/mineral content in fruits and veggies. On top of that, our soils have become so degraded from decades of modern Big Agriculture farming methods that the mineral and vitamin content in fruits and vegetables has dropped dramatically.
See, 70 years ago we could get most of our vitamins and minerals from eating small amounts of produce.
Today, that’s just not possible.
Even if you’re eating a much better diet than the Standard American Diet, the fact is even those eating healthfully have nutritional deficiencies or suboptimal intakes of many of these key nutrients.
Across the board, people’s bodies have been slowly stripped of the vital compounds they need to thrive.
And it’s absolutely hurting our health and energy levels in a massive way.
Get Your Energy Essentials & Superfoods Now!

Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.

Energy Essentials & Superfoods
$175.00
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Worldwide Shipping (FREE Shipping within the U.S.)*
* For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Worldwide Shipping (FREE Shipping within the U.S.)*
* For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
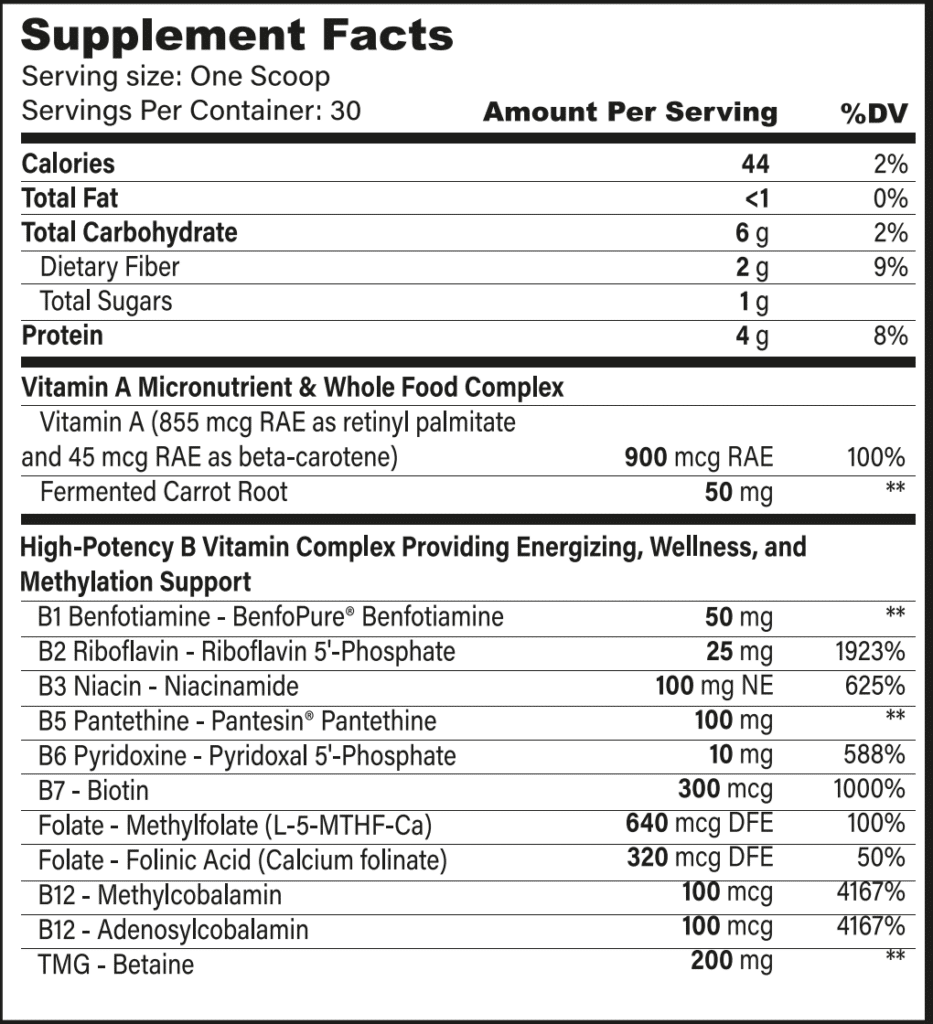
Energy Essentials & Superfoods Ingredients
Vitamin A Complex

Vitamin A refers to a group of fat-soluble retinoids and carotenoids, the main dietary ones being retinol and beta-carotene. Only retinol has biological activity within the body and can be considered “true” vitamin A. Beta-carotene can be converted by the body into vitamin A, but due to genetic differences, some people do this very poorly, so we’ve included retinol in its bioactive form in addition to beta-carotene.
Retinol is involved in regulating the growth and specialization of nearly every cell in the human body. In adults, it is critical for vision and immune function, with insufficient intakes leading to worsening eyesight (particularly at night) and a greater risk of infections and autoimmunity.1–3
We have the ability to make retinol from beta-carotene and a couple other carotenoids, which is why these precursors are considered a source of vitamin A. The problem is that the efficiency of the conversion process varies widely between individuals and populations due to genetic variations.4 The bioavailability of beta-carotene is also highly variable, depending on factors such as the food matrix, food preparation method, and one’s digestive and absorptive capacities.5 As such, beta-carotene is not a reliable source of vitamin A and we have provided most of one’s daily vitamin A requirement in the form of retinol.
B-Complex Vitamins

The B-complex vitamins include 8 different vitamins that are critical for methylation and energy production. Without them, we wouldn’t be able to turn the food we eat into cellular energy, our mitochondria couldn’t function, and we wouldn’t be able to turn the expression of certain genes on or off depending on our body’s needs.
- Thiamine (B1) is required to harvest energy from glucose, for the synthesis of energy-carrying molecules within mitochondria, and for nerve conduction.
- Riboflavin (B2) is the central unit for flavoproteins, which are necessary for mitochondrial energy production, antioxidant reactions and glutathione recycling, neurotransmitter regulation, and the metabolism of niacin, vitamin B6, and folate.
- Niacin (B3) is the central unit for NAD+ and NADP, which are required central to energy production, antioxidant reactions, detoxification, folate metabolism, the repair and regeneration of DNA and proteins, and the biosynthesis of fatty acids, cholesterol, and steroid hormones.
- Pantothenic acid (B5) is the central unit of coenzyme A, which plays a vital role in energy production and the metabolism of many nutrients, such as the breakdown of fats, carbohydrates, and proteins, the synthesis of fatty acids, ketones, cholesterol, steroid hormones, and neurotransmitters, genetic expression, and detoxification.
- Pyridoxal (B6) is required for amino acid metabolism, gene expression, and the breakdown of homocysteine.
- Biotin (B7) is the central component of several enzymes involved in energy production, amino acid metabolism, gene expression, and cell growth.
- Folate (B9) is required for methylation throughout our cells and mitochondria, DNA synthesis, cell division, the metabolism of glycine, and the breakdown of homocysteine.
- Cobalamin (B12) is necessary for just two important reactions in the body: (1) the breakdown of homocysteine alongside folate and, by virtue of this reaction, the recycling of folate, and (2) the breakdown of certain amino acids and fatty acids.
While all B-vitamin jobs are important, methylation and the breakdown of homocysteine are the ones that many people struggle with because of inadequate B-vitamin intakes and genetic mutations. Inappropriate methylation plays a role in many diseases, including cancer,6,7 neurodegeneration,8,9, and mood disorders like depression.10,11
Elevated levels of homocysteine is a major risk factor for cardiovascular diseases by directly causing blood vessel damage, reducing the flexibility of blood vessels, and altering processes involved in regulating blood fluidity.12 Additionally, high levels of homocysteine are strongly implicated in vascular dementia and impaired cognitive function for several reasons:13–16
- Neurotoxic
- Increases oxidative stress and inflammation
- Related to brain lesions and shrinkage
- Interferes with roper protein synthesis
Ensuring ample B-vitamin intake and status is a fundamental aspect of optimal health
Benfotiamine

Benfotiamine is a special fat-soluble form of thiamin that is capable of easily passing through cell membranes and providing free thiamine directly to the cell, particularly red blood cells and liver cells.17 Bioavailability studies have shown that benfotiamine increases serum thiamine concentrations 5- to 10-fold more than other forms of thiamine.18,19
Benfotiamine works to protect the body in several key ways:
- Activates an enzyme (transketolase) that reduces the creation of advanced glycation endproducts (AGEs) and protects against oxidative stress.25,26
- Increases cell survival and regeneration.27–29
- Reduces inflammation.30,31
- Improves brain health.32
Through these mechanisms, research has shown benfotiamine and thiamine supplementation to have notable health benefits, especially in those with diabetes:
- In healthy smokers, 1050 mg/d of benfotiamine was able to cut the smoking-induced impairment of endothelial function in half.33
- In type 2 diabetes, 1050 mg/d was able to completely prevent the 35% decline in blood vessel function after eating a meal rich in glycation endproducts.34<.sup>
- In type 2 diabetes, 150 mg/d of thiamine for 1 month was able to significantly reduce fasting glucose concentrations and improve leptin sensitivity.35
- Several studies have shown 200–600 mg/d of benfotiamine to improve diabetic neuropathy.36–39
- In adults with osteoarthritis, just 50 mg/d was able to reduce AGE concentrations, reduce pain, and improve physical function.40
- In Alzheimer’s disease, 300 mg/d for 18 months was able to improve cognitive function.41
Riboflavin 5' Phosphate

Riboflavin is critical for numerous functions in the body, but its most important role for most people is ensuring the proper function of the methylenetetrahydrofolate reductase (MTHFR) enzyme, which is necessary to recycle folate and detoxify homocysteine. A significant number of people have a mutation in MTHFR (the 677C→T variant) that reduces this enzymes activity, making them slow to convert 5,10-methylene-THF to 5-methyl-THF, the primary circulatory form of folate utilized in homocysteine detoxification.42
The mutation makes it more difficult for MTHFR to bind to a riboflavin-containing molecule (FAD) that is required for it to function.43,44 Supplementing riboflavin and saturating riboflavin stores in the body overcomes this reduced binding ability by having more FAD available to bind to, making the enzyme function normally again.
In other words, this mutation only causes a problem when riboflavin intake is inadequate. When you divide up people based on their riboflavin status, only those who are deficient in riboflavin have elevated homocysteine even in the presence of the MTHFR polymorphism.45
Although the dietary reference intake is set at 1–2 mg/d for healthy adults, this amount is not sufficient to saturate riboflavin stores for most people, particularly those with chronic inflammation that reduces riboflavin absorption.46
A meta-analysis of 14 studies found that supplementing with 2–5 mg/d above-and-beyond this reference intake leads to improved riboflavin status.47 Moreover, supplementing with just 2 mg/d has been shown to drop homocysteine by 40% in those with the MTHFR polymorphism.48
It’s also worth noting that riboflavin can lower blood pressure in those with the MTHFR polymorphism,49 which may be secondary to reducing homocysteine. Several interventions have noted significant reductions in blood pressure (5–10 mmHg) with less than 2 mg/d of riboflavin supplementation.50–52
Niacinamide

Niacinamide is a direct precurser to NAD+, which is involved in numerous notable health benefits like protecting against neurodegeneration, enhancing insulin sensitivity, increasing mitochondrial biogenesis, and sirtuin functions involved in “healthy aging”. 55,57
Pantesin

Pantesin (pantethine) is a form of pantothenic acid that not only satisfies our requirements but also has unique blood lipid–towering properties. A 2005 review of 28 small clinical trials with 64 participants with dyslipidemia reported a and 4 month reductions in LDL-C of 10% and 20% triglycerides of 14% and 33 % and increase in HDL-C of 6% and 8% respectively.58
Pyridoxal 5' Phosphate

Pyridoxal 5’ Phosphate (P5P) is the only biologically active form of vitamin B6, playing a vital role in the function of over 100 enzymes that catalyze essential chemical reactions in the human body. Adequate B6 is essential for various aspects of protein metabolism, the synthesis of glutathione from homocysteine via the transsulfuration pathway, and the metabolism of folate.
Biotin

Biotin (also known as vitamin H) is a key factor in helping our body convert food into energy. Biotin is also critical for hair, skin, and nail health.
Folate

A notable percentage of the population harbors variants in the methylenetetrahydrofolate reductase (MTHFR) enzyme, which is necessary to recycle folate and detoxify homocysteine. These variants reduce the ability of the MTHFR enzyme to turn 5,10-methylenetetrahydrofolate (5,10-methylene THF) into 5-methyl THF, which is the form required to recycle homocysteine.
Supplementing metafolin (5-methyl THF), bypasses this issue entirely, providing bioavailable folate for the body to use regardless of polymorphisms.
Betaine

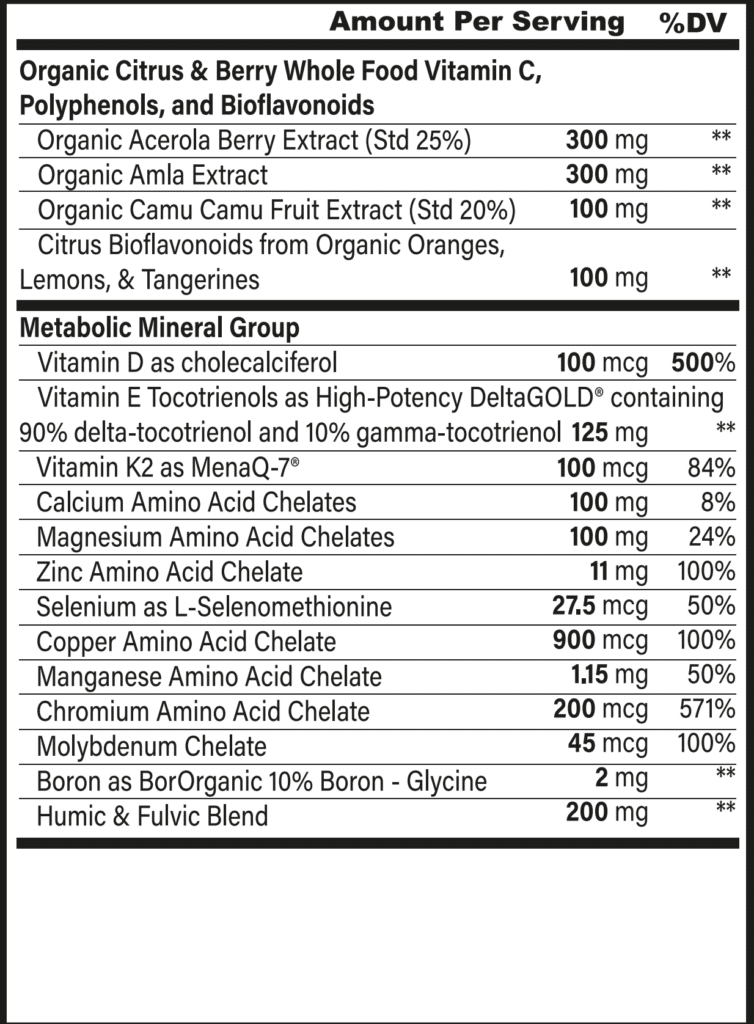
Vitamin D

Vitamin D is a unique nutrient because, although dietary sources of vitamin D do exist, our principle supply is through sunlight exposure. Making vitamin D wasn’t much of an issue throughout evolutionary history. A sunbathing adult produces the dietary equivalent of 10,000 to 25,000 IU when exposed to one minimal erythemal dose of UVB radiation, which is just enough to cause a slight pinkness to the skin 24 hours after exposure.
But things are a lot different today than they were 100, let alone 1 million years ago. The most obvious difference is that we spend most of our time indoors and away from sunlight. When we do go outside, we are wearing clothes that prevent sunlight from reaching the skin. When we go to the beach, we use sunscreen that blocks more than 99% of UVB radiation. This exposure problem becomes all the more ominous when we consider the role that skin pigmentation, geographical location, and genetics play in determining our ability to synthesize and utilize vitamin D.
Adding a little vitamin D into the diet can be a saving grace for those who cannot obtain sufficient sun exposure. While everyone will require different amounts, the Endocrine Society has recommended that people require 2,000 to 10,000 IU per day to achieve vitamin D sufficiency.112
Numerous meta-analyses have also linked low vitamin D levels to an increased risk of dying from any cause,113–117 cardiovascular diseases,113,114,116,118, and cancer.113,114,116,119,120
Vitamin E

Vitamin E is not one single chemical but instead refers to eight related fat-soluble compounds: four tocopherols and four tocotrienols. Most supplements use alpha-tocopherol as the source of vitamin E, but the tocotrienols have unique biologic functions not shared by the tocopherols.121
Gamma-tocopherol and delta-tocotrienol have emerged as vitamin E molecules with functions in health and disease that are clearly distinct from that of alpha-tocopherol.
Studies in humans suggest that tocotrienols possess incredibly potent antioxidant and anti-inflammatory effects that lend them benefits towards cardiovascular health, immune function, neuroprotection, and healthy aging. They have shown powerful benefits to preventing:122,123 124,125,126
- Cardiovascular disease (the #1 killer)
- Brain disease
- Cancer (including breast, colon, liver, lung, stomach, skin, pancreas, and prostate)
Metabolic Minerals

- Calcium is the major structural mineral of our bones and teeth and a critical messenger in cell-signaling pathways. When dietary calcium is inadequate, the body draws upon its reserves in the skeleton to supply what is needed.
- Magnesium is required for over 300 enzymes in the body to function properly, thereby regulating muscle contractions, blood pressure, insulin signaling, and the synthesis of DNA and proteins.
- Manganese plays an important role as both a constituent of enzymes and an activator of other enzyme’s activity, including those involved in antioxidant protection, metabolism, and bone development.
- Selenium exerts various biological effects as part of larger amino acid structures, with some of the best known being involved in glutathione and thyroid hormone production.
- Zinc plays an important role in regulating gene expression, ensuring the proper function of proteins, and catalyzing vital chemical reactions within the body.
- Copper is an essential cofactor for numerous redox reactions within energy production, iron metabolism, neurotransmission, and connective tissue development. It also serves as an essential component of the antioxidant enzyme, superoxide dismutase.
- Boron has key benefits for the skeletal system, anti-inflammatory benefits, dental health, and hormonal health (particularly testosterone and estrogen).180,182,183
- Molybdenum is a component of several enzymes within the body required for the metabolism of amino acids, DNA and RNA, and drugs.
- Chromium is critical for blood sugar management and correcting insulin resistance.
Fulvic and Humic Acid

Our formula also contains the wonder compounds fulvic and humic acid. Shilajit is a mixture of minerals used traditionally in Ayurveda, with the main bioactives being fulvic and humic acids. It’s believed to help transport other nutrients in the body, facilitate energy production within mitochondria, and supports the body’s detoxification processes.184,185 Clinical trials have shown that fulvic and humic acid supplementation can:
- Improve mitochondrial health and decrease fatigue (increases energy levels).
- Increase total and free testosterone levels in men.186,187
- Improve sperm parameters.187
- Binds and detoxifies heavy metals and other pollutants.
- Supports gut health.
Improves immune function. - Reduce oxidative stress.187
- Improves skin health.
- Improves brain health.
- Favorably alter gene expression in skeletal muscle for improved neuromuscular signaling and regeneration (faster recovery from workouts).188
- Enhance absorption of other compounds it is paired with.
Citrus Bioflavanoids

Citrus bioflavonoids are the phytonutrients found in citrus fruits, the most notable being hesperidin and naringenin. They are powerful antioxidant, immune-boosting, and anti-inflammatory compounds that have shown efficacy for preventing decreases in NAD+ that occur from metabolic dysfunction.68
Their anti-inflammatory effects have led to an intensive investigation for battling cancer and cardiovascular disease.69,70 Of course, part of this benefit comes from preventing mitochondrial dysfunction.71
To give an idea of the energy-boosting properties of citrus bioflavonoids, one study found that supplementation in combination with another citrus compound, synephrine, could increase metabolic rate by 18%.72
Acerola Berry Extract

Acerola berries are a rich source of vitamin C (1–4.5%) and phytochemicals.73,74 In fact, the vitamin C of acerola berries is more bioavailable and better retained in the body than vitamin C supplements.75
Research has implicated acerola in numerous beneficial aspects of our health:
- Protects against DNA damage.76–79
- Reduces fat cell dysfunction, oxidative stress, and low-grade inflammation that develop alongside obesity.78,80
- Promotes greater antioxidant potential within detoxification organs like the liver.81
- Increases brain mitochondrial function and energy production.82
Camu Camu

Amla

Amla is an Ayurvedic herb used traditionally for enhancing vitality and promoting longevity. It’s a potent supplier of vitamin C and bioactive phytochemicals that are able to promote mitochondrial biogenesis, up-regulate antioxidant systems, and facilitate energy production.95 These effects also lend it great therapeutic potential as a neuroprotective,96 anti-cancer,97 and general health agent.98
In particular, amla has incredible benefits towards blood glucose and blood lipids that have been documented in both healthy adults and adults with metabolic dysfunction (type 2 diabetes, metabolic syndrome, and dyslipidemia). The effects are comparable to those from diabetes drugs and statins.
In a study of non-diabetic and type 2 diabetic adults, 1–3 g/d of amla for 3 weeks reduced fasting blood glucose by 15–30% in non-diabetic adults and by 34–37% in those with type 2 diabetes, being as effective as the drug glibenclamide.99 It also lowered LDL-C and triglycerides, while HDL-C increased.
In healthy adults, taking 500 mg/d of amla for 4 weeks improved vascular fluidity, reduced an index for vascular age, and reduced markers of oxidative stress.100
A double-blind, randomized controlled trial using 1000 mg/d of amla for 12 weeks in dyslipidemic patients found that it reduced the atherogenic index by 39%, triglycerides by 15%, and LDL-C by 20%. 101
A randomized controlled trial comparing 500–1000 mg/d of amla with a statin in people with type 2 diabetes found that all were equivalently effective at improving endothelial function, increasing nitric oxide, reducing oxidative stress, and reducing inflammation.102 Amla was nearly as effective at improving blood lipids too.
In a study of individuals with GERD, 2000 mg/d of amla for 4 weeks significantly reduced the frequency and severity of heartburn.105
In a double-blind, randomized controlled trial comparing 500 and 1000 mg/d of amla for 12 weeks in individuals with metabolic syndrome, both doses similarly reduced oxidative stress (20–30%), increased glutathione concentrations (24–50%), increased nitric oxide (40–50%), and reduced inflammation (40–54%).103 It also positively affected blood lipids.
A clinical study of adults with dyslipidemia found that 500 mg/d of amla for 6 weeks was as effective as a statin at correcting the blood lipid abnormalities.104
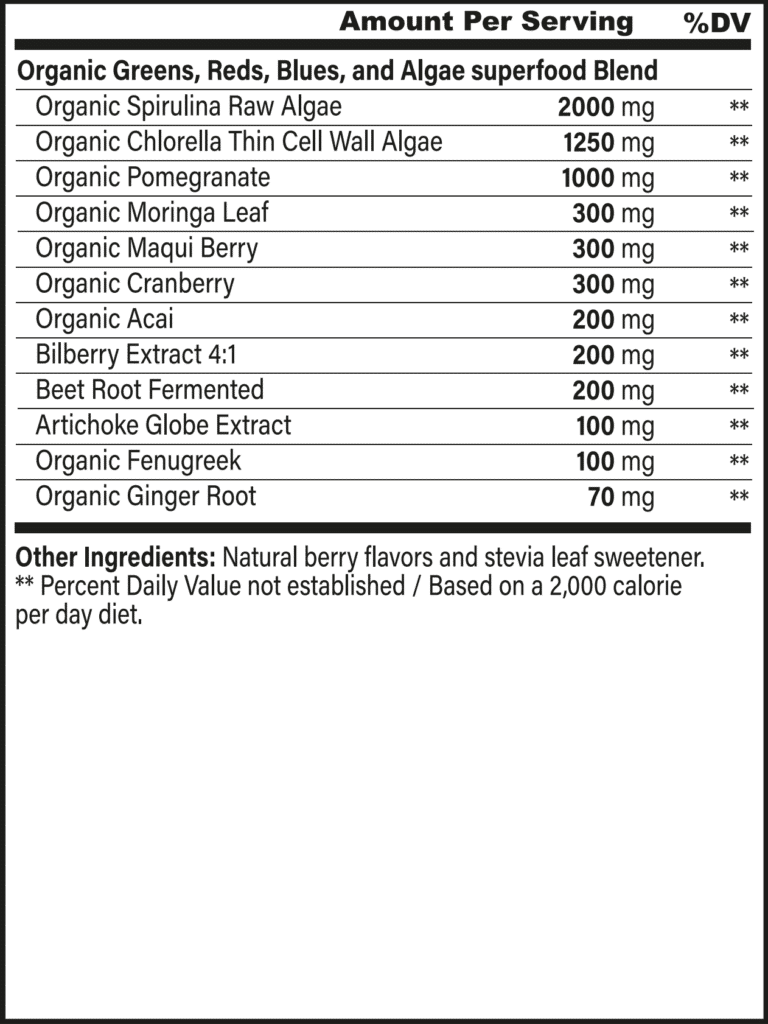
Chlorella Thin Cell Wall Algae

Chlorella is a microalga providing what’s called the Chlorella Growth Factor (CGF) — an extract composed of amino acids, peptides, proteins, vitamins, sugars, and nucleic acids.128
Studies have shown that chlorella has potent anti-inflammatory properties through activating the master anti-inflammatory regulator nuclear factor kappa B (NFκB).129 It also reduces levels of oxidative stress and increases markers of antioxidant activity.130
A meta-analysis of 19 randomized control trials with 797 participants revealed that Chlorella significantly decreases fasting blood glucose, blood pressure, and blood lipids.131 Additionally, it enhances immune function by stimulating the innate and adaptive immune soldiers.132 Other research has shown that it helps detoxify the body by enhancing the excretion of toxicants, including heavy metals.133,134 It’s an all-around amazing superfood for cardiovascular health, detoxification, and metabolic health.
Moringa Leaf

Moringa leaf is an important herb in Ayurvedic medicine, native to the Himalayan mountains and India, used to treat infections, high blood pressure, and diabetes. Research has demonstrated it has several beneficial health effects:135
- Reducing blood pressure and blood lipids
- Reducing inflammation and oxidative stress
- Reducing insulin resistance and blood glucose
- Reducing DNA damage and causing cancer cell death
- Protecting the liver
Spirulina

Spirulina is a group of cyanobacteria (not true algae as many believe) traditionally eaten by native African populations and the Aztecs of Central America.154 It is, by far, one of the most potent health supplements in existence thanks to a little molecule it contains called C-Phycocyanin.
This bioactive constituent of spirulina has powerful anti-inflammatory, antioxidant, antitumor, and immunostimulating properties thanks to its ability to inhibit NADPH oxidase activity,155,156 which serves as an important source of oxidative stress in many disease states.157,158 For example, it’s NADPH oxidase that drives the glucotoxicity and pancreatic damage seen in type 2 diabetes.159 Spirulina also protects against mitochondrial dysfunction and degeneration.160
The incredible health benefits of spirulina are evidenced in Gilbert’s Syndrome, a genetic condition where individuals have elevated levels of bilirubin,161 which is structurally similar to C-phycocyanin and also inhibits NADPH oxidase.162 People with Gilbert’s Syndrome have:
- A 50% lower risk of death from any cause compared to the general population.163
- Less oxidative stress.164
- Improved blood vessel function.164
- Lower rates of cardiovascular disease and mortality.165,166
- Longer telomeres.167
Ginger Root

Ginger is a culinary and medicinal herb in many cultures, with modern research confirming it has potent health benefits. While most people are familiar with its benefits for treating nausea, the effects extend far beyond that. Ginger supplementation has been shown to help battle:206,207
- Oxidative stress
- Inflammation
- Cancer
- Allergies
- Cognitive decline
- Diabetes
- Inflammatory bowel diseases
- Heart disease
- Dyslipidemia
Acai

Acai berries are a Brazilian superfruit native to the Amazon rainforest. They have an incredibly high amount of antioxidants, edging out other antioxidant-rich fruits like blueberries and cranberries.213,214 It’s also been shown to bolster mitochondrial function and energy production.215
Research has strongly suggested that acai berries hold promise for battling chronic diseases through reducing inflammation and oxidative stress:
- Battle cardiovascular disease through reducing blood lipid oxidation.216–218
- Fight cognitive decline with aging.219–222
- Protect against cancer and reduce its ability to spread and proliferate.223
- Reduce biomarkers of muscle damage following strenuous exercise, increase serum antioxidant status, and lower blood lipids.224
- Protects against liver damage and inflammation.225
- Prevent diabetes by improving insulin sensitivity and glycemic control.226–228
Bilberry Extract

Bilberries are a dark purple berry originating in Europe and closely related to blueberries. They possess a diverse array of anthocyanin phytochemicals similar to blueberries,229,230 which have potent antioxidant and anti-inflammatory effects.231
Regular intake could help fight cardiovascular disease, diabetes, and worsening vision.232,233
- Over just 3 weeks, supplementing with 300 mg/d of bilberry anthocyanins reduced inflammatory biomarkers by 38–60%, including suppressing the activation of the master inflammatory regulator, NF-kappaB.234 Similar observations have been made with bilberry juice,235 as well as fresh bilberries.236
- After just one dose of bilberry extract (470 mg), the glucose and insulin response to a glucose tolerance test was reduced by 18% each.237 Regular consumption of bilberries over 2 months improves fasting glucose and insulin secretion.238
- Over 2 months, consuming a mixture of berries, including bilberries, improved platelet function, blood pressure, and blood lipids.239 Supplementation with 320 mg bilberry anthocyanins reduced LDL by 14% and increased HDL by 11%.240
- In those with glaucoma, daily intake of 120 mg of bilberry anthocyanins for 2 years improved visual function by 30% compared to placebo, whose vision worsened.241
- In those with dry eyes, supplementing with 160 mg/d of bilberry extract for just one month improved dry eye symptoms by 55%.242
- In office workers suffering from eye strain, 160–480 mg/d of bilberry extract reduced eye fatigue caused by working with video display terminals.243–245
Maqui Berry

Maqui berries are an exotic, dark-purple fruit native to South America. They are a rich source of anthocyanins that possess strong antioxidant abilities,247,248 and ultimately provide up to three times more antioxidants than blackberries, blueberries, strawberries, or raspberries. 249,250
- Studies have shown that maqui berries could be a powerful ally against inflammatory diseases.251
- In particular, they have been shown to reduce inflammation of blood vessels,252 which could help prevent cardiovascular diseases.
- In smokers, 2 grams per day reduced markers of lung inflammation.253
- In a three-month clinical study in people with prediabetes, 180 mg of maqui berry extract once daily reduced average blood sugar levels by 5%, which was enough to bring their blood glucose levels back into normal range.254 It also reduced their LDL-c and increased their HDL-c.
- A small 30-day study in 13 people with dry eyes found that 30–60 mg of a concentrated maqui berry extract each day increased tear production by roughly 50%.255 Follow-up research demonstrated similar effects after just a month, along with reduced eye fatigue.256
Cranberry

Cranberries are best-known for their use in treating urinary tract infections. And they work. A meta-analysis of 10 studies found that cranberry juice reduces the likelihood of developing urinary tract infections,282 especially in women with recurrent episodes.283
These benefits are owed to the fact that cranberries are rich in A-type proanthocyanidins (PACs), in contrast to the B-type PACs present in most other fruits. Studies in humans and animals have shown cranberries to lower inflammation, increase nitric oxide synthesis, reduce blood pressure, manage blood lipids, improve endothelial function, and lower oxidative stress.284–286
- Drinking cranberry juice suppresses the growth of H. pylori infections in both adults and children after as little as 3 weeks.287,288
- Regular intake of cranberry juice (400–500 mL/d) increases HDL and decreases LDL,289,290 reduces LDL oxidation and the beginnings of atherosclerosis,291,292 lowers oxidative stress,293 and improves blood vessel function.294
- Cranberry phytochemicals prevent mitochondrial function, oxidative stress, and inflammation.295,296
Pomegranate

Pomegranate is a rich source of ellagitannins, potent antioxidants that can be further broken down into other antioxidant compounds like ellagic acid and urolithins.257,258 These substances have been heavily investigated for their cardiovascular, anti-cancer, and mitochondrial benefits.
Regular intake of pomegranate juice has been shown to reduce blood lipid oxidation and the accumulation of plaque in arteries over the course of 1–3 years,259 particularly in those who have higher levels of oxidative stress.260,261 Pomegranate effectively uses its potent antioxidant abilities to prevent the development of atherosclerosis.262,263
Additionally, pomegranate has been shown to help prevent and treat several types of cancer, including prostate, breast, lung, colon, and skin cancers.264,265 A phase II clinical trial in men with prostate cancer showed that regularly consuming pomegranate juice caused a 12% decrease in cell proliferation, a 17% increase in apoptosis, a 23% increase in serum nitric oxide, a reduction in oxidative stress, and a reduction in blood lipid oxidation.266
By far the most important benefit of pomegranates is their ability to stimulate mitochondrial function and mitophagy.267 Mitophagy (mitochondria + autophagy) is a quality control pathway that preserves mitochondrial health by targeting damaged mitochondria for autophagic degradation, making anything that facilitates mitophagy absolutely vital for optimal health and disease prevention.
Consuming pomegranate leads to the creation of urolithin A by our gut microbes, which is then absorbed into circulation where it induces mitophagy throughout the body and potentially extends lifespan in animal research.268 In particular, urolithin A is the single most potent activator of mitophagy (mitochondrial repair) in existence. And this compound is exclusively made from primarily ellagic acid in the diet, and pomegranate is one of the richest sources of ellagic acid. In fact, giving healthy adult humans urolithin A daily for one month improved markers of mitochondrial function!269
6 major things you should expect from your multivitamin...and what Energy Essentials & Superfoods delivers:6
SUPREME QUALITY
Carefully sourced, premium ingredients in the right ratios that will get you results. Taking Energy Essentials & Superfoods won't make you feel sick, and it won't leech nutrients from your body.
EFFECTIVE DOSAGES
Serving sizes that actually do something for your health. Feel more energetic and notice an overall greater sense of wellness after just a few daily doses. (Most supplement companies put in only 1/10th to 1/50th of the real effective doses. We are the only company making a product like this that actually uses real dose of the ingredients. For example, 1,500mg of spirulina instead of 50 mg, or 1,500 mg of pomegranate instead of 30 mg. Look at the label of ours and compare it to other brands to see for yourself.)
BIOAVAILABLE INGREDIENTS
Absorption is guaranteed, no matter the state of your gut health. Your body also knows how to receive each of the 45 vitamins, minerals, and superfoods in this multivitamin - at the same time. (No need to worry about competing nutrients.)
COMPLETE TRANSPARENCY
Zero hidden ingredients, fillers, junk, or harmful synthetics. What you see on the label is what you get - and what your body deserves.
RESEARCH-BASED FORMULATION
Pure science in every serving. No gimmicks. No marketing BS. Just mountains of science proving WHY you need all 45 ingredients in Energy Essentials & Superfoods.
MANUFACTURING BEST PRACTICES
Manufactured in the U.S. in an FDA-approved facility. Each bottle is independently lab-verified for purity, consistency, and quality. Delivered to your door exactly as it was created - a next-generation, ultra-premium multivitamin that's like nothing else out there.
Even if you don't think you need it.
The bad press around multivitamins is rampant, but it’s not entirely accurate. See below, where I bust some pretty big multivitamin myths. (And prove why Energy Essentials should be a key player in your supplement lineup.)
Even if you haven't taken a multivitamin in years.
Think you’re doing “just fine” without a high-quality supplement? Take another look at the long list of ways Energy Essentials & Superfoods can help you and see how much more your body can benefit when you add it to your daily routine. (There are 36 known benefits!)
Even if the one you're currently taking is "good enough".
I promise you, Energy Essentials & Superfoods is light years better than any other multivitamin you might be loyal to..Because, unlike just about every big name brand, each of the 45 ingredients in Energy Essentials & Superfoods are:
EASILY ABSORBED
No one wants to spend money on a supplement that moves right through the body, including us!
Enhanced absorption is KEY, so we’ve formulated Energy Essentials & Superfoods to include ONLY the most bioavailable forms of each of the 45 vitamins, minerals, botanicals, and potent antioxidants on the label.
You won’t find a single cheap or synthetic ingredient that could compromise absorption – or your health. (See for yourself – complete details below!)
And if you’re worried about certain ingredients ‘competing’ with each other – don’t be. When provided in their most bioavailable forms (in effective doses), your body is designed to absorb every nutrient in Energy Essentials & Superfoods. Yes – even when taken at the same time.
We’ve also included critical absorption enhancers like fulvic acid and humic acid to further optimize absorptions and shuttle these powerful nutrients directly into your cells.
IN EFFECTIVE DOSES
As is the case with our entire line of Energy Blueprint supplements, every dose of every ingredient in Energy Essentials & Superfoods is substantial enough to create positive change.
That means the recommended daily dose of Energy Essentials & Superfoods is robust.
While the serving size of most multivitamins is 1-3 grams, we suggest you take one 15-gram scoop of Energy Essentials. Every day.
Because here’s the hard truth — you simply cannot fit everything your body needs into ‘1-a-day’ or even 2 capsules (and anyone who tells you otherwise does NOT care about getting you results).
While that may seem like an overload, I promise you — every ingredient ratio is perfectly balanced and in line with exactly what your body needs to pump out abundant energy.
Not too much to do harm, not too little to do nothing. That’s the key. Our formulation is just right and is just what your body needs to fill in any nutrient gaps and optimize your health and energy.
SAFE TO TAKE
Whether your nutrient levels are already “normal” or need a bet of a boost, Energy Essentials is completely safe.*
There are zero synthetics or harmful ingredients that could harm you or make you feel ill.
And every single one of the superior quality vitamins, minerals, and superfoods in our premium formulation are:
- Sourced to the highest standards and independently lab certified
- Non GMO, gluten-free, soy-free, dairy-free
- Proven by research to have zero negative side effects
- Considered for long-term safety (no toxic build-up)
- Guaranteed to work WITH your body, not against it (no leaching of critical nutrients
BONUS: We’ve also carefully curated this stack of ingredients to minimize allergenic potential.
*Of course, consult your physician before taking EE if you are pregnant or have a pre-existing condition, or are taking any pharmaceuticals that could react negatively with EE.
What people say about Energy Essentials & Superfoods
Ari Whitten. I have only this week been able to start taking Energy Essentilas, have had it a while. OMG... only 3/4 a scoop. It is just amazing. I am now 71 and for the past week of taking this my energy has been better than ever. Clearly I was missing something that the this formula has because I feel sooooo much better. I have immense energy. Thank you so much."
Kristine G.
I was skeptical of your products... I finally got around to trying all three of your products and honestly...changed my life. Especially Energy Essentails. I can't express the difference I've noticed in my health with that."
Matthew
Get Your Energy Essentials & Superfoods Now!
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
$175.00
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Worldwide Shipping (FREE Shipping within the U.S.)*
* For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Energy Essentials & Superfoods
(FREE shipping within the U.S.)* *For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Worldwide Shipping (FREE Shipping within the U.S.)*
* For international orders: All shipments are shipped Duties, Customs, & Taxes unpaid. Buyer is responsible for delivery fees.
Reference
1. See, A. W.-M. & Clagett-Dame, M. The temporal requirement for vitamin A in the developing eye: mechanism of action in opticfissure closure and new roles for the vitamin in regulating cell proliferationand adhesion in the embryonic retina. Dev. Biol. 325, 94–105 (2009).
2. Green, H. N. & Mellanby, E. VITAMIN A AS ANANTI-INFECTIVE AGENT. Br. Med. J. 2, 691–696 (1928).
3. Raverdeau, M. & Mills, K. H. G. Modulation of Tcell and innate immune responses by retinoic Acid. J. Immunol. 192, 2953–2958(2014).
4. Borel, P. & Desmarchelier, C. Genetic VariationsAssociated with Vitamin A Status and Vitamin A Bioavailability. Nutrients 9,(2017).
5. Weber, D. & Grune, T. The contribution ofβ-carotene to vitamin A supply of humans. Mol. Nutr. Food Res. 56, 251–258(2012).
6. Wajed, S. A., Laird, P. W. & DeMeester, T. R. DNAmethylation: an alternative pathway to cancer. Ann. Surg. 234, 10–20 (2001).
7. Luczak, M. W. & Jagodziński, P. P. The role of DNAmethylation in cancer development. Folia Histochem. Cytobiol. 44, 143–154(2006).
8. Iraola-Guzmán, S., Estivill, X. & Rabionet, R. DNAmethylation in neurodegenerative disorders: a missing link between genome andenvironment? Clin. Genet. 80, 1–14 (2011).
9. Lu, H., Liu, X., Deng, Y. & Qing, H. DNAmethylation, a hand behind neurodegenerative diseases. Front. Aging Neurosci.5, 85 (2013).
10. Reynolds, E. H., Carney, M. W. & Toone, B. K.Methylation and mood. Lancet 2, 196–198 (1984).
11. Bottiglieri, T. Folate, vitamin B12, andneuropsychiatric disorders. Nutr. Rev. 54, 382–390 (1996).
12. Ganguly, P. & Alam, S. F. Role of homocysteine inthe development of cardiovascular disease. Nutr. J. 14, 6 (2015).
13. Price, B. R., Wilcock, D. M. & Weekman, E. M.Hyperhomocysteinemia as a Risk Factor for Vascular Contributions to CognitiveImpairment and Dementia. Front. Aging Neurosci. 10, 350 (2018).
14. West, R. K. et al. Homocysteine and cognitive functionin very elderly nondemented subjects. Am. J. Geriatr. Psychiatry 19, 673–677(2011).
15. Garcia, A. & Zanibbi, K. Homocysteine andcognitive function in elderly people. CMAJ 171, 897–904 (2004).
16. Moretti, R. & Caruso, P. The Controversial Role ofHomocysteine in Neurology: From Labs to Clinical Practice. Int. J. Mol. Sci.20, (2019).
17. Wada, T. et al. A new thiamine derivative,S-benzoylthiamine O-monophosphate. Science 134, 195–196 (1961).
18. Loew, D. Pharmacokinetics of thiamine derivativesespecially of benfotiamine. Int. J. Clin. Pharmacol. Ther. 34, 47–50 (1996).
19. Xie, F. et al. Pharmacokinetic study of benfotiamineand the bioavailability assessment compared to thiamine hydrochloride. J. Clin.Pharmacol. 54, 688–695 (2014).
20. Pácal, L., Kuricová, K. & Kaňková, K. Evidence foraltered thiamine metabolism in diabetes: Is there a potential to oppose gluco-and lipotoxicity by rational supplementation? World J. Diabetes 5, 288–295(2014).
21. Jain, A., Mehta, R., Al-Ani, M., Hill, J. A. &Winchester, D. E. Determining the Role of Thiamine Deficiency in Systolic HeartFailure: A Meta-Analysis and Systematic Review. J. Card. Fail. 21, 1000–1007(2015).
22. Page, G. L. J., Laight, D. & Cummings, M. H.Thiamine deficiency in diabetes mellitus and the impact of thiamine replacementon glucose metabolism and vascular disease. Int. J. Clin. Pract. 65, 684–690(2011).
23. Luong, K. V. Q. & Nguyen, L. T. H. The impact ofthiamine treatment in the diabetes mellitus. J. Clin. Med. Res. 4, 153–160(2012).
24. Thornalley, P. J. et al. High prevalence of low plasmathiamine concentration in diabetes linked to a marker of vascular disease.Diabetologia 50, 2164–2170 (2007).
25. Hammes, H.-P. et al. Benfotiamine blocks three majorpathways of hyperglycemic damage and prevents experimental diabeticretinopathy. Nat. Med. 9, 294–299 (2003).
26. Kihm, L. P. et al. Benfotiamine protects againstperitoneal and kidney damage in peritoneal dialysis. J. Am. Soc. Nephrol. 22,914–926 (2011).
27. Gadau, S. et al. Benfotiamine accelerates the healingof ischaemic diabetic limbs in mice through protein kinase B/Akt-mediatedpotentiation of angiogenesis and inhibition of apoptosis. Diabetologia 49,405–420 (2006).
28. Beltramo, E., Nizheradze, K., Berrone, E., Tarallo, S.& Porta, M. Thiamine and benfotiamine prevent apoptosis induced by highglucose-conditioned extracellular matrix in human retinal pericytes. Diabetes.Metab. Res. Rev. 25, 647–656 (2009).
29. Verma, S., Reddy, K. & Balakumar, P. The defensiveeffect of benfotiamine in sodium arsenite-induced experimental vascularendothelial dysfunction. Biol. Trace Elem. Res. 137, 96–109 (2010).
30. Bozic, I. et al. Benfotiamine attenuates inflammatoryresponse in LPS stimulated BV-2 microglia. PLoS One 10, e0118372 (2015).
31. Yadav, U. C. S., Kalariya, N. M., Srivastava, S. K.& Ramana, K. V. Protective role of benfotiamine, a fat-soluble vitamin B1analogue, in lipopolysaccharide-induced cytotoxic signals in murinemacrophages. Free Radic. Biol. Med. 48, 1423–1434 (2010).
32. Sun, X.-J. et al. Benfotiamine prevents increasedβ-amyloid production in HEK cells induced by high glucose. Neurosci. Bull. 28,561–566 (2012).
33. Stirban, A. et al. Benfotiamine counteractssmoking-induced vascular dysfunction in healthy smokers. Int. J. Vasc. Med.2012, 968761 (2012).
34. Stirban, A. et al. Benfotiamine prevents macro- andmicrovascular endothelial dysfunction and oxidative stress following a mealrich in advanced glycation end products in individuals with type 2 diabetes.Diabetes Care 29, 2064–2071 (2006).
35. González-Ortiz, M., Martínez-Abundis, E.,Robles-Cervantes, J. A., Ramírez-Ramírez, V. & Ramos-Zavala, M. G. Effectof thiamine administration on metabolic profile, cytokines and inflammatorymarkers in drug-naïve patients with type 2 diabetes. Eur. J. Nutr. 50, 145–149(2011).
36. Stracke, H., Gaus, W., Achenbach, U., Federlin, K.& Bretzel, R. G. Benfotiamine in diabetic polyneuropathy (BENDIP): resultsof a randomised, double blind, placebo-controlled clinical study. Exp. Clin.Endocrinol. Diabetes 116, 600–605 (2008).
37. Winkler, G. et al. Effectiveness of differentbenfotiamine dosage regimens in the treatment of painful diabetic neuropathy.Arzneimittelforschung 49, 220–224 (1999).
38. Haupt, E., Ledermann, H. & Köpcke, W. Benfotiaminein the treatment of diabetic polyneuropathy–a three-week randomized, controlledpilot study (BEDIP study). Int. J. Clin. Pharmacol. Ther. 43, 71–77 (2005).
39. Nikolić, A., Kacar, A., Lavrnić, D., Basta, I. &Apostolski, S. [The effect of benfothiamine in the therapy of diabeticpolyneuropathy]. Srp. Arh. Celok. Lek. 137, 594–600 (2009).
40. Garg, S., Syngle, A. & Vohra, K. Efficacy andtolerability of advanced glycation end-products inhibitor in osteoarthritis: arandomized, double-blind, placebo-controlled study. Clin. J. Pain 29, 717–724(2013).
41. Pan, X. et al. Long-Term Cognitive Improvement AfterBenfotiamine Administration in Patients with Alzheimer’s Disease. Neurosci.Bull. 32, 591–596 (2016).
42. Leclerc, D., Sibani, S. & Rozen, R. Molecularbiology of methylenetetrahydrofolate reductase (MTHFR) and overview ofmutations/polymorphisms. in MTHFR polymorphisms and disease 15–34 (CRC Press,2005).
43. Guenther, B. D. et al. The structure and properties ofmethylenetetrahydrofolate reductase from Escherichia coli suggest how folateameliorates human hyperhomocysteinemia. Nat. Struct. Biol. 6, 359–365 (1999).
44. Yamada, K., Chen, Z., Rozen, R. & Matthews, R. G.Effects of common polymorphisms on the properties of recombinant humanmethylenetetrahydrofolate reductase. Proc. Natl. Acad. Sci. U. S. A. 98,14853–14858 (2001).
45. García-Minguillán, C. J. et al. Riboflavin statusmodifies the effects of methylenetetrahydrofolate reductase (MTHFR) andmethionine synthase reductase (MTRR) polymorphisms on homocysteine. Genes Nutr.9, 435 (2014).
46. Anandam, K. Y. et al. Effect of the proinflammatorycytokine TNF-α on intestinal riboflavin uptake: inhibition mediated viatranscriptional mechanism(s). Am. J. Physiol. Cell Physiol. 315, C653–C663(2018).
47. Hoey, L., McNulty, H. & Strain, J. J. Studies ofbiomarker responses to intervention with riboflavin: a systematic review. Am.J. Clin. Nutr. 89, 1960S–1980S (2009).
48. McNulty, H. et al. Riboflavin lowers homocysteine inindividuals homozygous for the MTHFR 677C->T polymorphism. Circulation 113,74–80 (2006).
49. McNulty, H., Strain, J. J., Hughes, C. F. & Ward,M. Riboflavin, MTHFR genotype and blood pressure: A personalized approach toprevention and treatment of hypertension. Mol. Aspects Med. 53, 2–9 (2017).
50. Horigan, G. et al. Riboflavin lowers blood pressure incardiovascular disease patients homozygous for the 677C–>T polymorphism inMTHFR. J. Hypertens. 28, 478–486 (2010).
51. Wilson, C. P. et al. Riboflavin offers a targetedstrategy for managing hypertension in patients with the MTHFR 677TT genotype: a4-y follow-up. Am. J. Clin. Nutr. 95, 766–772 (2012).
52. Wilson, C. P. et al. Blood pressure in treatedhypertensive individuals with the MTHFR 677TT genotype is responsive tointervention with riboflavin: findings of a targeted randomized trial.Hypertension 61, 1302–1308 (2013).
53. Zempleni, J., Galloway, J. R. & McCormick, D. B.Pharmacokinetics of orally and intravenously administered riboflavin in healthyhumans. Am. J. Clin. Nutr. 63, 54–66 (1996).
54. Madigan, S. M. et al. Riboflavin and vitamin B-6intakes and status and biochemical response to riboflavin supplementation infree-living elderly people. Am. J. Clin. Nutr. 68, 389–395 (1998).
55. Braidy, N. et al. Role of Nicotinamide AdenineDinucleotide and Related Precursors as Therapeutic Targets for Age-RelatedDegenerative Diseases: Rationale, Biochemistry, Pharmacokinetics, and Outcomes.Antioxid. Redox Signal. 30, 251–294 (2019).
56. Rajman, L., Chwalek, K. & Sinclair, D. A.Therapeutic Potential of NAD-Boosting Molecules: The In Vivo Evidence. CellMetab. 27, 529–547 (2018).
57. Johnson, S. & Imai, S.-I. NAD + biosynthesis,aging, and disease. F1000Res. 7, 132 (2018).
58. McRae, M. P. Treatment of hyperlipoproteinemia withpantethine: A review and analysis of efficacy and tolerability. Nutr. Res. 25,319–333 (2005).
59. Steenge, G. R., Verhoef, P. & Katan, M. B. Betainesupplementation lowers plasma homocysteine in healthy men and women. J. Nutr.133, 1291–1295 (2003).
60. Olthof, M. R., van Vliet, T., Boelsma, E. &Verhoef, P. Low dose betaine supplementation leads to immediate and long termlowering of plasma homocysteine in healthy men and women. J. Nutr. 133,4135–4138 (2003).
61. Rajaie, S. & Esmaillzadeh, A. Dietary choline andbetaine intakes and risk of cardiovascular diseases: review of epidemiologicalevidence. ARYA Atheroscler. 7, 78–86 (2011).
62. Zhao, G. et al. Betaine in Inflammation: MechanisticAspects and Applications. Front. Immunol. 9, 1070 (2018).
63. Eussen, S. J. P. M. et al. The association of betaine,homocysteine and related metabolites with cognitive function in Dutch elderlypeople. Br. J. Nutr. 98, 960–968 (2007).
64. Knight, L. S., Piibe, Q., Lambie, I., Perkins, C.& Yancey, P. H. Betaine in the Brain: Characterization of Betaine Uptake,its Influence on Other Osmolytes and its Potential Role in Neuroprotection fromOsmotic Stress. Neurochem. Res. 42, 3490–3503 (2017).
65. Kim, S. J., Lee, L., Kim, J. H., Lee, T. H. &Shim, I. Antidepressant-like effects of lycii radicis cortex and betaine in theforced swimming test in rats. Biomol. Ther. 21, 79–83 (2013).
66. Di Pierro, F., Orsi, R. & Settembre, R. Role ofbetaine in improving the antidepressant effect of S-adenosyl-methionine inpatients with mild-to-moderate depression. J. Multidiscip. Healthc. 8, 39–45(2015).
67. Di Pierro, F. & Settembre, R. Preliminary resultsof a randomized controlled trial carried out with a fixed combination ofS-adenosyl-L-methionine and betaine versus amitriptyline in patients with milddepression. Int. J. Gen. Med. 8, 73–78 (2015).
68. Boesten, D. M. P. H. J., von Ungern-Sternberg, S. N.I., den Hartog, G. J. M. & Bast, A. Protective Pleiotropic Effect ofFlavonoids on NAD+ Levels in Endothelial Cells Exposed to High Glucose. Oxid.Med. Cell. Longev. 2015, 894597 (2015).
69. Benavente-García, O. & Castillo, J. Update on usesand properties of citrus flavonoids: new findings in anticancer,cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 56,6185–6205 (2008).
70. Tejada, S. et al. Potential Anti-inflammatory Effectsof Hesperidin from the Genus Citrus. Curr. Med. Chem. 25, 4929–4945 (2018).
71. Kumar, A., Prakash, A. & Dogra, S. Naringinalleviates cognitive impairment, mitochondrial dysfunction and oxidative stressinduced by D-galactose in mice. Food Chem. Toxicol. 48, 626–632 (2010).
72. Stohs, S. J. et al. Effects of p-synephrine alone andin combination with selected bioflavonoids on resting metabolism, bloodpressure, heart rate and self-reported mood changes. Int. J. Med. Sci. 8,295–301 (2011).
73. Prakash, A. & Baskaran, R. Acerola, an untappedfunctional superfruit: a review on latest frontiers. J. Food Sci. Technol. 55,3373–3384 (2018).
74. Cardoso, P. C., Tomazini, A. P. B., Stringheta, P. C.,Ribeiro, S. M. R. & Pinheiro-Sant’Ana, H. M. Vitamin C and carotenoids inorganic and conventional fruits grown in Brazil. Food Chem. 126, 411–416(2011).
75. Uchida, E. et al. Absorption and excretion of ascorbicacid alone and in acerola (Malpighia emarginata) juice: comparison in healthyJapanese subjects. Biol. Pharm. Bull. 34, 1744–1747 (2011).
76. Nunes, R. da S. et al. Antigenotoxicity andantioxidant activity of Acerola fruit (Malpighia glabra L.) at two stages ofripeness. Plant Foods Hum. Nutr. 66, 129–135 (2011).
77. Nunes, R. D. S. et al. Genotoxic and antigenotoxicactivity of acerola (Malpighia glabra L.) extract in relation to the geographicorigin. Phytother. Res. 27, 1495–1501 (2013).
78. Leffa, D. D. et al. Corrective effects of acerola(Malpighia emarginata DC.) juice intake on biochemical and genotoxicalparameters in mice fed on a high-fat diet. Mutat. Res. 770, 144–152 (2014).
79. Horta, R. N. et al. Protective effects of acerolajuice on genotoxicity induced by iron in vivo. Genet. Mol. Biol. 39, 122–128(2016).
80. Dias, F. M. et al. Acerola (Malpighia emarginata DC.)juice intake protects against alterations to proteins involved in inflammatoryand lipolysis pathways in the adipose tissue of obese mice fed a cafeteriadiet. Lipids Health Dis. 13, 24 (2014).
81. Leffa, D. D. et al. Effects of Supplemental AcerolaJuice on the Mineral Concentrations in Liver and Kidney Tissue Samples of MiceFed with Cafeteria Diet. Biol. Trace Elem. Res. 167, 70–76 (2015).
82. Leffa, D. D. et al. Effects of Acerola (Malpighiaemarginata DC.) Juice Intake on Brain Energy Metabolism of Mice Fed a CafeteriaDiet. Mol. Neurobiol. 54, 954–963 (2017).
83. Justi, K. C., Visentainer, J. V., Evelázio de Souza,N. & Matsushita, M. Nutritional composition and vitamin C stability instored camu-camu (Myrciaria dubia) pulp. Arch. Latinoam. Nutr. 50, 405–408(2000).
84. Fracassetti, D., Costa, C., Moulay, L. &Tomás-Barberán, F. A. Ellagic acid derivatives, ellagitannins,proanthocyanidins and other phenolics, vitamin C and antioxidant capacity oftwo powder products from camu-camu fruit (Myrciaria dubia). Food Chem. 139,578–588 (2013).
85. Langley, P. C., Pergolizzi, J. V., Jr, Taylor, R., Jr& Ridgway, C. Antioxidant and associated capacities of Camu camu (Myrciariadubia): a systematic review. J. Altern. Complement. Med. 21, 8–14 (2015).
86. Fidelis, M. et al. Camu-camu seed (Myrciaria dubia) –From side stream to anantioxidant, antihyperglycemic, antiproliferative,antimicrobial, antihemolytic, anti-inflammatory, and antihypertensiveingredient. Food Chem. 310, 125909 (2020).
87. De Souza Schmidt Gonçalves, A. E., Lajolo, F. M. &Genovese, M. I. Chemical composition and antioxidant/antidiabetic potential ofBrazilian native fruits and commercial frozen pulps. J. Agric. Food Chem. 58,4666–4674 (2010).
88. Genovese, M. I., Da Silva Pinto, M., De Souza SchmidtGonçalves, A. E. & Lajolo, F. M. Bioactive Compounds and AntioxidantCapacity of Exotic Fruits and Commercial Frozen Pulps from Brazil. Food Sci.Technol. Int. 14, 207–214 (2008).
89. Neri-Numa, I. A., Soriano Sancho, R. A., Pereira, A.P. A. & Pastore, G. M. Small Brazilian wild fruits: Nutrients, bioactivecompounds, health-promotion properties and commercial interest. Food Res. Int.103, 345–360 (2018).
90. Nascimento, O. V., Boleti, A. P. A., Yuyama, L. K. O.& Lima, E. S. Effects of diet supplementation with Camu-camu (Myrciariadubia HBK McVaugh) fruit in a rat model of diet-induced obesity. An. Acad.Bras. Cienc. 85, 355–363 (2013).
91. Akachi, T. et al. 1-methylmalate from camu-camu(Myrciaria dubia) suppressed D-galactosamine-induced liver injury in rats.Biosci. Biotechnol. Biochem. 74, 573–578 (2010).
92. Inoue, T., Komoda, H., Uchida, T. & Node, K.Tropical fruit camu-camu (Myrciaria dubia) has anti-oxidative andanti-inflammatory properties. J. Cardiol. 52, 127–132 (2008).
93. Balisteiro, D. M., Araujo, R. L. de, Giacaglia, L. R.& Genovese, M. I. Effect of clarified Brazilian native fruit juices onpostprandial glycemia in healthy subjects. Food Res. Int. 100, 196–203 (2017).
94. Miyashita, T. et al. Data on a single oral dose ofcamu camu (Myrciaria dubia) pericarp extract on flow-mediated vasodilation andblood pressure in young adult humans. Data Brief 16, 993–999 (2018).
95. Yamamoto, H. et al. Amla Enhances Mitochondrial SpareRespiratory Capacity by Increasing Mitochondrial Biogenesis and AntioxidantSystems in a Murine Skeletal Muscle Cell Line. Oxid. Med. Cell. Longev. 2016,1735841 (2016).
96. Husain, I. et al. Exploring the multifacetedneuroprotective actions of Emblica officinalis (Amla): a review. Metab. BrainDis. 34, 957–965 (2019).
97. Baliga, M. S. & Dsouza, J. J. Amla (Emblicaofficinalis Gaertn), a wonder berry in the treatment and prevention of cancer.Eur. J. Cancer Prev. 20, 225–239 (2011).
98. Krishnaveni, M. & Mirunalini, S. Therapeuticpotential of Phyllanthus emblica (amla): the ayurvedic wonder. J. Basic Clin.Physiol. Pharmacol. 21, 93–105 (2010).
99. Akhtar, M. S., Ramzan, A., Ali, A. & Ahmad, M.Effect of Amla fruit (Emblica officinalis Gaertn.) on blood glucose and lipidprofile of normal subjects and type 2 diabetic patients. Int. J. Food Sci.Nutr. 62, 609–616 (2011).
100. Kapoor, M. P., Suzuki, K., Derek, T., Ozeki, M. &Okubo, T. Clinical evaluation of Emblica Officinalis Gatertn (Amla) in healthyhuman subjects: Health benefits and safety results from a randomized,double-blind, crossover placebo-controlled study. Contemp Clin Trials Commun17, 100499 (2020).
101. Upadya, H. et al. A randomized, double blind, placebocontrolled, multicenter clinical trial to assess the efficacy and safety ofEmblica officinalis extract in patients with dyslipidemia. BMC Complement.Altern. Med. 19, 27 (2019).
102. Usharani, P., Fatima, N. & Muralidhar, N. Effectsof Phyllanthus emblica extract on endothelial dysfunction and biomarkers ofoxidative stress in patients with type 2 diabetes mellitus: a randomized,double-blind, controlled study. Diabetes Metab. Syndr. Obes. 6, 275–284 (2013).
103. Usharani, P., Merugu, P. L. & Nutalapati, C.Evaluation of the effects of a standardized aqueous extract of Phyllanthusemblica fruits on endothelial dysfunction, oxidative stress, systemicinflammation and lipid profile in subjects with metabolic syndrome: arandomised, double blind, placebo controlled clinical study. BMC Complement.Altern. Med. 19, 97 (2019).
104. Gopa, B., Bhatt, J. & Hemavathi, K. G. Acomparative clinical study of hypolipidemic efficacy of Amla (Emblica officinalis)with 3-hydroxy-3-methylglutaryl-coenzyme-A reductase inhibitor simvastatin.Indian J. Pharmacol. 44, 238–242 (2012).
105. Karkon Varnosfaderani, S. et al. Efficacy and safetyof Amla (Phyllanthus emblica L.) in non-erosive reflux disease: a double-blind,randomized, placebo-controlled clinical trial. J. Integr. Med. 16, 126–131(2018).
106. Hosseinzadeh, H. & Nassiri-Asl, M. Review of theprotective effects of rutin on the metabolic function as an important dietaryflavonoid. J. Endocrinol. Invest. 37, 783–788 (2014).
107. Al-Dhabi, N. A., Arasu, M. V., Park, C. H. & Park,S. U. An up-to-date review of rutin and its biological and pharmacologicalactivities. EXCLI J. 14, 59–63 (2015).
108. Ganeshpurkar, A. & Saluja, A. K. ThePharmacological Potential of Rutin. Saudi Pharm J 25, 149–164 (2017).
109. Enogieru, A. B., Haylett, W., Hiss, D. C., Bardien, S.& Ekpo, O. E. Rutin as a Potent Antioxidant: Implications forNeurodegenerative Disorders. Oxid. Med. Cell. Longev. 2018, 6241017 (2018).
110. Ragheb, S. R., El Wakeel, L. M., Nasr, M. S. &Sabri, N. A. Impact of Rutin and Vitamin C combination on oxidative stress andglycemic control in patients with type 2 diabetes. Clin Nutr ESPEN 35, 128–135(2020).
111. Sattanathan, K., Dhanapal, C. K., Umarani, R. &Manavalan, R. Beneficial health effects of rutin supplementation in patientswith diabetes mellitus. J. Basic Appl. Pharm. Sci. 1, 227 (2011).
112. Holick, M. F. et al. Evaluation, treatment, andprevention of vitamin D deficiency: an Endocrine Society clinical practiceguideline. J. Clin. Endocrinol. Metab. 96, 1911–1930 (2011).
113. Gaksch, M. et al. Vitamin D and mortality: Individualparticipant data meta-analysis of standardized 25-hydroxyvitamin D in 26916individuals from a European consortium. PLoS One 12, e0170791 (2017).
114. Schöttker, B. et al. Vitamin D and mortality:meta-analysis of individual participant data from a large consortium of cohortstudies from Europe and the United States. BMJ 348, g3656 (2014).
115. Garland, C. F. et al. Meta-analysis of all-causemortality according to serum 25-hydroxyvitamin D. Am. J. Public Health 104,e43–50 (2014).
116. Chowdhury, R. et al. Vitamin D and risk of causespecific death: systematic review and meta-analysis of observational cohort andrandomised intervention studies. BMJ 348, g1903 (2014).
117. Aspelund, T. et al. Effect of Genetically Low25-Hydroxyvitamin D on Mortality Risk: Mendelian Randomization Analysis in 3Large European Cohorts. Nutrients 11, (2019).
118. Gholami, F. et al. The association between circulating25-hydroxyvitamin D and cardiovascular diseases: a meta-analysis of prospectivecohort studies. BMC Cardiovasc. Disord. 19, 248 (2019).
119. Zhang,Y. et al. Association between vitamin D supplementation and mortality:systematic review and meta-analysis. BMJ 366, l4673 (2019).
120. Han, J. et al. 25-Hydroxyvitamin D and Total CancerIncidence and Mortality: A Meta-Analysis of Prospective Cohort Studies.Nutrients 11, (2019).
121. Sen, C. K., Khanna, S. & Roy, S. Tocotrienols:Vitamin E beyond tocopherols. Life Sci. 78, 2088–2098 (2006).
122. Sen, C. K., Khanna, S. & Roy, S. Tocotrienol: thenatural vitamin E to defend the nervous system? Ann. N. Y. Acad. Sci. 1031,127–142 (2004).
123. Sylvester, P. W., Kaddoumi, A., Nazzal, S. & ElSayed, K. A. The value of tocotrienols in the prevention and treatment ofcancer. J. Am. Coll. Nutr. 29, 324S–333S (2010).
124. Aggarwal, B. B., Sundaram, C., Prasad, S. &Kannappan, R. Tocotrienols, the vitamin E of the 21st century: its potentialagainst cancer and other chronic diseases. Biochem. Pharmacol. 80, 1613–1631(2010).
125. Sen, C. K., Khanna, S., Rink, C. & Roy, S.Tocotrienols: the emerging face of natural vitamin E. Vitam. Horm. 76, 203–261(2007).
126. Meganathan, P. & Fu, J.-Y. Biological Propertiesof Tocotrienols: Evidence in Human Studies. Int. J. Mol. Sci. 17, (2016).
127. Ben Salem, M. et al. Pharmacological Studies ofArtichoke Leaf Extract and Their Health Benefits. Plant Foods Hum. Nutr. 70,441–453 (2015).
128. Merchant, R. E. & Andre, C. A. A review of recentclinical trials of the nutritional supplement Chlorella pyrenoidosa in the treatmentof fibromyalgia, hypertension, and ulcerative colitis. Altern. Ther. HealthMed. 7, 79–91 (2001).
129. Lee, H.-S., Choi, C.-Y., Cho, C. & Song, Y.Attenuating effect of chlorella supplementation on oxidative stress andNFkappaB activation in peritoneal macrophages and liver of C57BL/6 mice fed onan atherogenic diet. Biosci. Biotechnol. Biochem. 67, 2083–2090 (2003).
130. Panahi, Y. et al. Investigation of the effects ofChlorella vulgaris supplementation on the modulation of oxidative stress inapparently healthy smokers. Clin. Lab. 59, 579–587 (2013).
131. Fallah, A. A. et al. Effect of Chlorellasupplementation on cardiovascular risk factors: A meta-analysis of randomizedcontrolled trials. Clin. Nutr. 37, 1892–1901 (2018).
132. Kwak, J. H. et al. Beneficial immunostimulatory effectof short-term Chlorella supplementation: enhancement of natural killer cellactivity and early inflammatory response (randomized, double-blinded,placebo-controlled trial). Nutr. J. 11, 53 (2012).
133. Lee, I. et al. Detoxification of chlorella supplementon heterocyclic amines in Korean young adults. Environ. Toxicol. Pharmacol. 39,441–446 (2015).
134. Uchikawa, T. et al. Enhanced elimination of tissuemethylmercury in Parachlorella beijerinckii-fed mice. J. Toxicol. Sci. 36,121–126 (2011).
135. Vergara-Jimenez, M., Almatrafi, M. M. & Fernandez,M. L. Bioactive Components in Moringa Oleifera Leaves Protect against ChronicDisease. Antioxidants (Basel) 6, (2017).
136. Lee, D., Nishizawa, M., Shimizu, Y. & Saeki, H.Anti-inflammatory effects of dulse (Palmaria palmata) resulting from thesimultaneous water-extraction of phycobiliproteins and chlorophyll a. Food Res.Int. 100, 514–521 (2017).
137. Millan-Linares, M. C. et al. Nutraceutical Extractfrom Dulse (Palmaria palmata L.) Inhibits Primary Human Neutrophil Activation.Mar. Drugs 17, (2019).
138. Mendez, R. et al. Antiobesogenic Potential of SeaweedDulse (Palmaria palmata) in High-fat Fed C57BL/6 J Mice (P21-014-19). Curr DevNutr 3, (2019).
139. Kim, J. K. & Park, S. U. Current potential healthbenefits of sulforaphane. EXCLI J. 15, 571–577 (2016).
140. de Figueiredo, S. M., Binda, N. S., Nogueira-Machado,J. A., Vieira-Filho, S. A. & Caligiorne, R. B. The antioxidant propertiesof organosulfur compounds (sulforaphane). Recent Pat. Endocr. Metab. ImmuneDrug Discov. 9, 24–39 (2015).
141. Zeng, Y. et al. Preventive and Therapeutic Role ofFunctional Ingredients of Barley Grass for Chronic Diseases in Human Beings.Oxid. Med. Cell. Longev. 2018, 3232080 (2018).
142. Yu, Y.-M., Chang, W.-C., Chang, C.-T., Hsieh, C.-L.& Tsai, C. E. Effects of young barley leaf extract and antioxidativevitamins on LDL oxidation and free radical scavenging activities in type 2diabetes. Diabetes Metab. 28, 107–114 (2002).
143. Wirngo, F. E., Lambert, M. N. & Jeppesen, P. B.The Physiological Effects of Dandelion (Taraxacum Officinale) in Type 2Diabetes. Rev. Diabet. Stud. 13, 113–131 (2016).
144. Ma, C. et al. Anti-inflammatory effects of waterextract of Taraxacum mongolicum hand.-Mazz on lipopolysaccharide-inducedinflammation in acute lung injury by suppressing PI3K/Akt/mTOR signalingpathway. J. Ethnopharmacol. 168, 349–355 (2015).
145. Hu, G. et al. Effects of aqueous extracts of TaraxacumOfficinale on expression of tumor necrosis factor-alpha and intracellularadhesion molecule 1 in LPS-stimulated RMMVECs. BMC Complement. Altern. Med. 17,38 (2017).
146. Park, C. M., Cho, C. W. & Song, Y. S. TOP 1 and 2,polysaccharides from Taraxacum officinale, inhibit NFκB-mediated inflammationand accelerate Nrf2-induced antioxidative potential through the modulation ofPI3K-Akt signaling pathway in RAW 264.7 cells. Food Chem. Toxicol. 66, 56–64(2014).
147. Koh, Y.-J., Cha, D.-S., Ko, J.-S., Park, H.-J. &Choi, H.-D. Anti-inflammatory effect of Taraxacum officinale leaves onlipopolysaccharide-induced inflammatory responses in RAW 264.7 cells. J. Med.Food 13, 870–878 (2010).
148. Ovadje, P., Ammar, S., Guerrero, J.-A., Arnason, J. T.& Pandey, S. Dandelion root extract affects colorectal cancer proliferationand survival through the activation of multiple death signalling pathways.Oncotarget 7, 73080–73100 (2016).
149. Ovadje, P., Hamm, C. & Pandey, S. Efficientinduction of extrinsic cell death by dandelion root extract in human chronicmyelomonocytic leukemia (CMML) cells. PLoS One 7, e30604 (2012).
150. Koo, H.-N. et al. Taraxacum officinale inducescytotoxicity through TNF-alpha and IL-1alpha secretion in Hep G2 cells. LifeSci. 74, 1149–1157 (2004).
151. Menghini, L. et al. Antiproliferative, protective andantioxidant effects of artichoke, dandelion, turmeric and rosemary extracts andtheir formulation. Int. J. Immunopathol. Pharmacol. 23, 601–610 (2010).
152. Lau, D. & Plotkin, B. J. Antimicrobial and biofilmeffects of herbs used in traditional Chinese medicine. Nat. Prod. Commun. 8,1617–1620 (2013).
153. Clare, B. A., Conroy, R. S. & Spelman, K. Thediuretic effect in human subjects of an extract of Taraxacum officinale foliumover a single day. J. Altern. Complement. Med. 15, 929–934 (2009).
154. Ciferri, O. Spirulina, the edible microorganism.Microbiol. Rev. 47, 551–578 (1983).
155. Liu, Q., Huang, Y., Zhang, R., Cai, T. & Cai, Y.Medical Application of Spirulina platensis Derived C-Phycocyanin. Evid. Based.Complement. Alternat. Med. 2016, 7803846 (2016).
156. McCarty, M. F. Clinical potential of Spirulina as asource of phycocyanobilin. J. Med. Food 10, 566–570 (2007).
157. Panday, A., Sahoo, M. K., Osorio, D. & Batra, S.NADPH oxidases: an overview from structure to innate immunity-associatedpathologies. Cell. Mol. Immunol. 12, 5–23 (2015).
158. Bedard, K. & Krause, K.-H. The NOX family ofROS-generating NADPH oxidases: physiology and pathophysiology. Physiol. Rev.87, 245–313 (2007).
159. McCarty, M. F., Barroso-Aranda, J. & Contreras, F.NADPH oxidase mediates glucolipotoxicity-induced beta cell dysfunction–clinicalimplications. Med. Hypotheses 74, 596–600 (2010).
160. Nawrocka, D., Kornicka, K., Śmieszek, A. & Marycz,K. Spirulina platensis Improves Mitochondrial Function Impaired by ElevatedOxidative Stress in Adipose-Derived Mesenchymal Stromal Cells (ASCs) andIntestinal Epithelial Cells (IECs), and Enhances Insulin Sensitivity in EquineMetabolic Syndrome (EMS) Horses. Mar. Drugs 15, (2017).
161. Olsson, R. & Stigendal, L. Clinical experiencewith isolated hyperbilirubinemia. Scand. J. Gastroenterol. 24, 617–622 (1989).
162. Hinds, T. D., Jr & Stec, D. E. Bilirubin, aCardiometabolic Signaling Molecule. Hypertension 72, 788–795 (2018).
163. Horsfall, L. J., Nazareth, I., Pereira, S. P. &Petersen, I. Gilbert’s syndrome and the risk of death: a population-basedcohort study. J. Gastroenterol. Hepatol. 28, 1643–1647 (2013).
164. Maruhashi, T. et al. Hyperbilirubinemia, augmentationof endothelial function, and decrease in oxidative stress in Gilbert syndrome.Circulation 126, 598–603 (2012).
165. Kundur, A. R., Singh, I. & Bulmer, A. C.Bilirubin, platelet activation and heart disease: a missing link tocardiovascular protection in Gilbert’s syndrome? Atherosclerosis 239, 73–84(2015).
166. Goel, A. & Aggarwal, R. Unconjugatedhyperbilirubinemia: a blessing in disguise? Journal of gastroenterology andhepatology vol. 28 1687–1689 (2013).
167. Tosevska, A. et al. Longer telomeres in chronic,moderate, unconjugated hyperbilirubinaemia: insights from a human study onGilbert’s Syndrome. Sci. Rep. 6, 22300 (2016).
180. Naghii, M. R., Mofid, M., Asgari, A. R., Hedayati, M.& Daneshpour, M.-S. Comparative effects of daily and weekly boronsupplementation on plasma steroid hormones and proinflammatory cytokines. J.Trace Elem. Med. Biol. 25, 54–58 (2011).
181. Newnham, R. E. Essentiality of boron for healthy bonesand joints. Environ. Health Perspect. 102 Suppl 7, 83–85 (1994).
182. Naghii, M. R. & Samman, S. The effect of boronsupplementation on its urinary excretion and selected cardiovascular riskfactors in healthy male subjects. Biol. Trace Elem. Res. 56, 273–286 (1997).
183. Nielsen, F. H., Hunt, C. D., Mullen, L. M. & Hunt,J. R. Effect of dietary boron on mineral, estrogen, and testosterone metabolismin postmenopausal women. FASEB J. 1, 394–397 (1987).
184. Meena, H., Pandey, H. K., Arya, M. C. & Ahmed, Z.Shilajit: A panacea for high-altitude problems. Int. J. Ayurveda Res. 1, 37–40(2010).
185. Agarwal, S. P., Khanna, R., Karmarkar, R., Anwer, M.K. & Khar, R. K. Shilajit: a review. Phytother. Res. 21, 401–405 (2007).
186. Pandit, S. et al. Clinical evaluation of purified Shilajiton testosterone levels in healthy volunteers. Andrologia 48, 570–575 (2016).
187. Biswas, T. K. et al. Clinical evaluation ofspermatogenic activity of processed Shilajit in oligospermia. Andrologia 42,48–56 (2010).
188. Das, A. et al. The Human Skeletal Muscle Transcriptomein Response to Oral Shilajit Supplementation. J. Med. Food 19, 701–709 (2016).
189. Burge, K., Gunasekaran, A., Eckert, J. & Chaaban,H. Curcumin and Intestinal Inflammatory Diseases: Molecular Mechanisms ofProtection. Int. J. Mol. Sci. 20, (2019).
190. Ng, Q. X. et al. A Meta-Analysis of the Clinical Useof Curcumin for Irritable Bowel Syndrome (IBS). J. Clin. Med. Res. 7, (2018).
191. Ghosh, S. S., He, H., Wang, J., Gehr, T. W. &Ghosh, S. Curcumin-mediated regulation of intestinal barrier function: Themechanism underlying its beneficial effects. Tissue Barriers 6, e1425085(2018).
192. Peterson, C. T. et al. Effects of Turmeric andCurcumin Dietary Supplementation on Human Gut Microbiota: A Double-Blind,Randomized, Placebo-Controlled Pilot Study. J Evid Based Integr Med 23,2515690X18790725 (2018).
193. Di Meo, F., Margarucci, S., Galderisi, U., Crispi, S.& Peluso, G. Curcumin, Gut Microbiota, and Neuroprotection. Nutrients 11,(2019).
194. Hewlings, S. J. & Kalman, D. S. Curcumin: A Reviewof Its’ Effects on Human Health. Foods 6, (2017).
195. Soto-Urquieta, M. G. et al. Curcumin restoresmitochondrial functions and decreases lipid peroxidation in liver and kidneysof diabetic db/db mice. Biol. Res. 47, 74 (2014).
196. Ahmad, A., Alghamdi, S. S., Mahmood, K. & Afzal,M. Fenugreek a multipurpose crop: Potentialities and improvements. Saudi J.Biol. Sci. 23, 300–310 (2016).
197. Dawid-Pać, R. Medicinal plants used in treatment ofinflammatory skin diseases. Postepy Dermatol Alergol 30, 170–177 (2013).
198. Steels, E., Rao, A. & Vitetta, L. Physiologicalaspects of male libido enhanced by standardized Trigonella foenum-graecumextract and mineral formulation. Phytother. Res. 25, 1294–1300 (2011).
199. Rao, A., Steels, E., Inder, W. J., Abraham, S. &Vitetta, L. Testofen, a specialised Trigonella foenum-graecum seed extractreduces age-related symptoms of androgen decrease, increases testosteronelevels and improves sexual function in healthy aging males in a double-blindrandomised clinical study. Aging Male 19, 134–142 (2016).
200. Maheshwari, A. et al. Efficacy of FurosapTM, a novelTrigonella foenum-graecum seed extract, in Enhancing Testosterone Level andImproving Sperm Profile in Male Volunteers. Int. J. Med. Sci. 14, 58–66 (2017).
201. Wankhede, S., Mohan, V. & Thakurdesai, P.Beneficial effects of fenugreek glycoside supplementation in male subjectsduring resistance training: A randomized controlled pilot study. J Sport HealthSci 5, 176–182 (2016).
202. Rao, A., Steels, E., Beccaria, G., Inder, W. J. &Vitetta, L. Influence of a Specialized Trigonella foenum-graecum Seed Extract(Libifem), on Testosterone, Estradiol and Sexual Function in HealthyMenstruating Women, a Randomised Placebo Controlled Study. Phytother. Res. 29,1123–1130 (2015).
203. Kassaian, N., Azadbakht, L., Forghani, B. & Amini,M. Effect of fenugreek seeds on blood glucose and lipid profiles in type 2diabetic patients. Int. J. Vitam. Nutr. Res. 79, 34–39 (2009).
204. Sharma, R. D., Raghuram, T. C. & Rao, N. S. Effectof fenugreek seeds on blood glucose and serum lipids in type I diabetes. Eur.J. Clin. Nutr. 44, 301–306 (1990).
205. Kiss, R. et al. Insulin-Sensitizer Effects ofFenugreek Seeds in Parallel with Changes in Plasma MCH Levels in Healthy Volunteers.Int. J. Mol. Sci. 19, (2018).
207. Bode, A. M. & Dong, Z. Chapter 7 The Amazing andMighty Ginger. Herbal Medicine: Biomolecular and Clinical Aspects.
208. Mazidi, M., Gao, H.-K., Rezaie, P. & Ferns, G. A.The effect of ginger supplementation on serum C-reactive protein, lipid profileand glycaemia: a systematic review and meta-analysis. Food Nutr. Res. 60, 32613(2016).
209. Kanner, J., Harel, S. & Granit, R. Betalains–a newclass of dietary cationized antioxidants. J. Agric. Food Chem. 49, 5178–5185(2001).
210. Clifford, T., Howatson, G., West, D. J. &Stevenson, E. J. The potential benefits of red beetroot supplementation inhealth and disease. Nutrients 7, 2801–2822 (2015).
211. Bondonno, C. P., Croft, K. D. & Hodgson, J. M.Dietary Nitrate, Nitric Oxide, and Cardiovascular Health. Crit. Rev. Food Sci.Nutr. 56, 2036–2052 (2016).
212. Bondonno, C. P. et al. Vegetable-derived bioactivenitrate and cardiovascular health. Mol. Aspects Med. 61, 83–91 (2018).
213. Vanderpool, R. & Gladwin, M. T. Harnessing thenitrate-nitrite-nitric oxide pathway for therapy of heart failure withpreserved ejection fraction. Circulation vol. 131 334–336 (2015).
214. USDA Database for the Oxygen Radical AbsorbanceCapacity (ORAC) of Selected Foods, Release 2.
215. Lichtenthäler, R. et al. Total oxidant scavenging capacitiesof Euterpe oleracea Mart. (Açaí) fruits. Int. J. Food Sci. Nutr. 56, 53–64(2005).
216. Machado, A. K. et al. Neuroprotective Effects of Açaí(Euterpe oleracea Mart.) against Rotenone In Vitro Exposure. Oxid. Med. Cell.Longev. 2016, 8940850 (2016).
217. Feio, C. A. et al. Euterpe oleracea (açai) modifiessterol metabolism and attenuates experimentally-induced atherosclerosis. J.Atheroscler. Thromb. 19, 237–245 (2012).
218. de Souza, M. O. et al. The hypocholesterolemicactivity of açaí (Euterpe oleracea Mart.) is mediated by the enhancedexpression of the ATP-binding cassette, subfamily G transporters 5 and 8 andlow-density lipoprotein receptor genes in the rat. Nutr. Res. 32, 976–984(2012).
219. de Souza, M. O., Silva, M., Silva, M. E., Oliveira, R.de P. & Pedrosa, M. L. Diet supplementation with acai (Euterpe oleraceaMart.) pulp improves biomarkers of oxidative stress and the serum lipid profilein rats. Nutrition 26, 804–810 (2010).
220. Poulose, S. M. et al. Restoration of stressor-inducedcalcium dysregulation and autophagy inhibition by polyphenol-rich açaí (Euterpespp.) fruit pulp extracts in rodent brain cells in vitro. Nutrition 30, 853–862(2014).
221. Poulose, S. M., Bielinski, D. F., Carey, A., Schauss,A. G. & Shukitt-Hale, B. Modulation of oxidative stress, inflammation,autophagy and expression of Nrf2 in hippocampus and frontal cortex of rats fedwith açaí-enriched diets. Nutr. Neurosci. 20, 305–315 (2017).
222. Poulose, S. M. et al. Anthocyanin-rich açai (Euterpeoleracea Mart.) fruit pulp fractions attenuate inflammatory stress signaling inmouse brain BV-2 microglial cells. J. Agric. Food Chem. 60, 1084–1093 (2012).
223. Carey, A. N. et al. Dietary supplementation with thepolyphenol-rich açaí pulps (Euterpe oleracea Mart. and Euterpe precatoriaMart.) improves cognition in aged rats and attenuates inflammatory signaling inBV-2 microglial cells. Nutr. Neurosci. 20, 238–245 (2017).
224. Alessandra-Perini, J., Rodrigues-Baptista, K. C.,Machado, D. E., Nasciutti, L. E. & Perini, J. A. Anticancer potential,molecular mechanisms and toxicity of Euterpe oleracea extract (açaí): Asystematic review. PLoS One 13, e0200101 (2018).
225. Sadowska-Krępa, E. et al. Effects of supplementationwith acai (Euterpe oleracea Mart.) berry-based juice blend on the bloodantioxidant defence capacity and lipid profile in junior hurdlers. A pilotstudy. Biol. Sport 32, 161–168 (2015).
226. de Freitas Carvalho, M. M. et al. Effects of açai onoxidative stress, ER stress, and inflammation-related parameters in mice withhigh fat diet-fed induced NAFLD. Sci. Rep. 9, 8107 (2019).
227. Udani, J. K., Singh, B. B., Singh, V. J. &Barrett, M. L. Effects of Açai (Euterpe oleracea Mart.) berry preparation onmetabolic parameters in a healthy overweight population: a pilot study. Nutr.J. 10, 45 (2011).
228. Solverson, P. M. et al. An Anthocyanin-RichMixed-Berry Intervention May Improve Insulin Sensitivity in a Randomized Trialof Overweight and Obese Adults. Nutrients 11, (2019).
229. Belwal, T., Nabavi, S. F., Nabavi, S. M. &Habtemariam, S. Dietary Anthocyanins and Insulin Resistance: When Food Becomesa Medicine. Nutrients 9, (2017).
230. Burdulis, D., Ivanauskas, L., Dirse, V., Kazlauskas,S. & Razukas, A. Study of diversity of anthocyanin composition in bilberry(Vaccinium myrtillus L.) fruits. Medicina 43, 971–977 (2007).
231. Lätti, A. K., Riihinen, K. R. & Kainulainen, P. S.Analysis of anthocyanin variation in wild populations of bilberry (Vacciniummyrtillus L.) in Finland. J. Agric. Food Chem. 56, 190–196 (2008).
232. Colak, N. et al. Bog bilberry phenolics, antioxidantcapacity and nutrient profile. Food Chem. 201, 339–349 (2016).
233. Chu, W.-K., Cheung, S. C. M., Lau, R. A. W. &Benzie, I. F. F. Bilberry (Vaccinium myrtillus L.). in Herbal Medicine:Biomolecular and Clinical Aspects (eds. Benzie, I. F. F. & Wachtel-Galor,S.) (CRC Press/Taylor & Francis, 2012).
234. Chan, S. W. & Tomlinson, B. Effects of BilberrySupplementation on Metabolic and Cardiovascular Disease Risk. Molecules 25,(2020).
235. Karlsen, A. et al. Anthocyanins inhibit nuclearfactor-kappaB activation in monocytes and reduce plasma concentrations ofpro-inflammatory mediators in healthy adults. J. Nutr. 137, 1951–1954 (2007).
236. Karlsen, A. et al. Bilberry juice modulates plasmaconcentration of NF-kappaB related inflammatory markers in subjects atincreased risk of CVD. Eur. J. Nutr. 49, 345–355 (2010).
237. Kolehmainen, M. et al. Bilberries reduce low-gradeinflammation in individuals with features of metabolic syndrome. Mol. Nutr.Food Res. 56, 1501–1510 (2012).
238. Hoggard, N. et al. A single supplement of astandardised bilberry (Vaccinium myrtillus L.) extract (36 % wet weightanthocyanins) modifies glycaemic response in individuals with type 2 diabetescontrolled by diet and lifestyle. J. Nutr. Sci. 2, e22 (2013).
239. de Mello, V. D. et al. Fasting serum hippuric acid iselevated after bilberry (Vaccinium myrtillus) consumption and associates withimprovement of fasting glucose levels and insulin secretion in persons at highrisk of developing type 2 diabetes. Mol. Nutr. Food Res. 61, (2017).
240. Erlund, I. et al. Favorable effects of berryconsumption on platelet function, blood pressure, and HDL cholesterol. Am. J.Clin. Nutr. 87, 323–331 (2008).
241. Qin, Y. et al. Anthocyanin supplementation improvesserum LDL- and HDL-cholesterol concentrations associated with the inhibition ofcholesteryl ester transfer protein in dyslipidemic subjects. Am. J. Clin. Nutr.90, 485–492 (2009).
242. Shim, S. H., Kim, J. M., Choi, C. Y., Kim, C. Y. &Park, K. H. Ginkgo biloba extract and bilberry anthocyanins improve visualfunction in patients with normal tension glaucoma. J. Med. Food 15, 818–823(2012).
243. Riva, A. et al. The effect of a natural, standardizedbilberry extract (Mirtoselect®) in dry eye: a randomized, double blinded,placebo-controlled trial. Eur. Rev. Med. Pharmacol. Sci. 21, 2518–2525 (2017).
244. Liang, T. & Others. Effect of a Bilberry Extract(BILBERON®)—containing Diet on the Improvement of Eye Fatigue—related Symptoms(Ⅱ)―A Randomized, Double—blind, Placebo—controlled, Parallel—group ComparisonStudy―. 薬理と治療45, 1523–1534 (2017).
245. Ozawa, Y. et al. Bilberry extract supplementation forpreventing eye fatigue in video display terminal workers. J. Nutr. Health Aging19, 548–554 (2015).
246. Kosehira, M., Machida, N. & Kitaichi, N. A12-Week-Long Intake of Bilberry Extract (Vaccinium myrtillus L.) ImprovedObjective Findings of Ciliary Muscle Contraction of the Eye: A Randomized,Double-Blind, Placebo-Controlled, Parallel-Group Comparison Trial. Nutrients12, (2020).
247. Ooe, E. et al. Bilberry extract and anthocyaninssuppress unfolded protein response induced by exposure to blue LED light ofcells in photoreceptor cell line. Mol. Vis. 24, 621–632 (2018).
248. Quispe-Fuentes, I., Vega-Gálvez, A. & Aranda, M.Evaluation of phenolic profiles and antioxidant capacity of maqui (Aristoteliachilensis) berries and their relationships to drying methods. J. Sci. FoodAgric. 98, 4168–4176 (2018).
249. Davinelli, S., Bertoglio, J. C., Zarrelli, A., Pina,R. & Scapagnini, G. A Randomized Clinical Trial Evaluating the Efficacy ofan Anthocyanin-Maqui Berry Extract (Delphinol®) on Oxidative Stress Biomarkers.J. Am. Coll. Nutr. 34 Suppl 1, 28–33 (2015).
250. Schreckinger, M. E., Lotton, J., Lila, M. A. & deMejia, E. G. Berries from South America: a comprehensive review on chemistry,health potential, and commercialization. J. Med. Food 13, 233–246 (2010).
251. Araya, H., Clavijo, C. & Herrera, C. [Antioxidantcapacity of fruits and vegetables cultivated in Chile]. Arch. Latinoam. Nutr.56, 361–365 (2006).
252. Cespedes, C. L. et al. The chilean superfruitblack-berry Aristotelia chilensis (Elaeocarpaceae), Maqui as mediator ininflammation-associated disorders. Food Chem. Toxicol. 108, 438–450 (2017).
253. Watson, R. R. & Schönlau, F. Nutraceutical andantioxidant effects of a delphinidin-rich maqui berry extract Delphinol®: areview. Minerva Cardioangiol. 63, 1–12 (2015).
254. Vergara, D. et al. The intake of maqui (Aristoteliachilensis) berry extract normalizes H2O2 and IL-6 concentrations in exhaledbreath condensate from healthy smokers – an explorative study. Nutr. J. 14, 27(2015).
255. Alvarado, J., Schoenlau, F., Leschot, A., Salgad, A.M. & Vigil Portales, P. Delphinol® standardized maqui berry extractsignificantly lowers blood glucose and improves blood lipid profile inprediabetic individuals in three-month clinical trial. Panminerva Med. 58, 1–6(2016).
256. Hitoe, S., Tanaka, J. & Shimoda, H. MaquiBrightTMstandardized maqui berry extract significantly increases tear fluid productionand ameliorates dry eye-related symptoms in a clinical pilot trial. PanminervaMed. 56, 1–6 (2014).
257. Yamashita, S.-I., Suzuki, N., Yamamoto, K., Iio, S.-I.& Yamada, T. Effects of MaquiBright® on improving eye dryness and fatiguein humans: A randomized, double-blind, placebo-controlled trial. Afr. J.Tradit. Complement. Altern. Med. 9, 172–178 (2019).
258. Heber, D. Pomegranate Ellagitannins. in HerbalMedicine: Biomolecular and Clinical Aspects (eds. Benzie, I. F. F. &Wachtel-Galor, S.) (CRC Press/Taylor & Francis, 2012).
259. Ismail, T. et al. Ellagitannins in CancerChemoprevention and Therapy. Toxins 8, (2016).
260. Aviram, M. et al. Pomegranate juice consumption for 3years by patients with carotid artery stenosis reduces common carotidintima-media thickness, blood pressure and LDL oxidation. Clin. Nutr. 23,423–433 (2004).
261. Davidson, M. H. et al. Effects of consumption ofpomegranate juice on carotid intima-media thickness in men and women atmoderate risk for coronary heart disease. Am. J. Cardiol. 104, 936–942 (2009).
262. Basu, A., Newman, E. D., Bryant, A. L., Lyons, T. J.& Betts, N. M. Pomegranate polyphenols lower lipid peroxidation in adultswith type 2 diabetes but have no effects in healthy volunteers: a pilot study.J. Nutr. Metab. 2013, 708381 (2013).
263. Aviram, M. et al. Pomegranate juice flavonoids inhibitlow-density lipoprotein oxidation and cardiovascular diseases: studies inatherosclerotic mice and in humans. Drugs Exp. Clin. Res. 28, 49–62 (2002).
264. Rosenblat, M., Volkova, N., Attias, J., Mahamid, R.& Aviram, M. Consumption of polyphenolic-rich beverages (mostly pomegranateand black currant juices) by healthy subjects for a short term increased serumantioxidant status, and the serum’s ability to attenuate macrophage cholesterolaccumulation. Food Funct. 1, 99–109 (2010).
265. Zarfeshany, A., Asgary, S. & Javanmard, S. H.Potent health effects of pomegranate. Adv. Biomed. Res. 3, 100 (2014).
266. Sharma, P., McClees, S. F. & Afaq, F. Pomegranatefor Prevention and Treatment of Cancer: An Update. Molecules 22, (2017).
267. Pantuck, A. J. et al. Phase II study of pomegranatejuice for men with rising prostate-specific antigen following surgery orradiation for prostate cancer. Clin. Cancer Res. 12, 4018–4026 (2006).
268. Tan, S. et al. Pomegranate activates TFEB to promoteautophagy-lysosomal fitness and mitophagy. Sci. Rep. 9, 727 (2019).
269. Ryu, D. et al. Urolithin A induces mitophagy andprolongs lifespan in C. elegans and increases muscle function in rodents. Nat.Med. 22, 879–888 (2016).
270. Andreux, P. A. et al. The mitophagy activatorurolithin A is safe and induces a molecular signature of improved mitochondrialand cellular health in humans. Nat Metab 1, 595–603 (2019).
271. Tan, L. et al. Investigation on the Role of BDNF inthe Benefits of Blueberry Extracts for the Improvement of Learning and Memoryin Alzheimer’s Disease Mouse Model. J. Alzheimers. Dis. 56, 629–640 (2017).
272. Rendeiro, C. et al. Blueberry supplementation inducesspatial memory improvements and region-specific regulation of hippocampal BDNFmRNA expression in young rats. Psychopharmacology 223, 319–330 (2012).
273. Bensalem, J. et al. Polyphenol-rich extract from grapeand blueberry attenuates cognitive decline and improves neuronal function inaged mice. J. Nutr. Sci. 7, e19 (2018).
274. Winter, A. N. & Bickford, P. C. Anthocyanins andTheir Metabolites as Therapeutic Agents for Neurodegenerative Disease.Antioxidants (Basel) 8, (2019).
275. Krikorian, R. et al. Blueberry supplementationimproves memory in older adults. J. Agric. Food Chem. 58, 3996–4000 (2010).
276. Bowtell, J. L., Aboo-Bakkar, Z., Conway, M. E., Adlam,A.-L. R. & Fulford, J. Enhanced task-related brain activation and restingperfusion in healthy older adults after chronic blueberry supplementation.Appl. Physiol. Nutr. Metab. 42, 773–779 (2017).
277. Boespflug, E. L. et al. Enhanced neural activationwith blueberry supplementation in mild cognitive impairment. Nutr. Neurosci.21, 297–305 (2018).
278. Whyte, A. R., Cheng, N., Fromentin, E. & Williams,C. M. A Randomized, Double-Blinded, Placebo-Controlled Study to Compare the Safetyand Efficacy of Low Dose Enhanced Wild Blueberry Powder and Wild BlueberryExtract (ThinkBlueTM) in Maintenance of Episodic and Working Memory in OlderAdults. Nutrients 10, (2018).
279. Bergland, A. K. et al. Effects of AnthocyaninSupplementation on Serum Lipids, Glucose, Markers of Inflammation and Cognitionin Adults With Increased Risk of Dementia – A Pilot Study. Front. Genet. 10,536 (2019).
280. Whyte, A. R., Cheng, N., Butler, L. T., Lamport, D. J.& Williams, C. M. Flavonoid-Rich Mixed Berries Maintain and ImproveCognitive Function Over a 6 h Period in Young Healthy Adults. Nutrients 11,(2019).
281. Kent, K., Charlton, K. E., Netzel, M. & Fanning,K. Food-based anthocyanin intake and cognitive outcomes in human interventiontrials: a systematic review. J. Hum. Nutr. Diet. 30, 260–274 (2017).
282. Ma, L., Sun, Z., Zeng, Y., Luo, M. & Yang, J.Molecular Mechanism and Health Role of Functional Ingredients in Blueberry forChronic Disease in Human Beings. Int. J. Mol. Sci. 19, (2018).
283. Jepson, R. G. & Craig, J. C. Cranberries forpreventing urinary tract infections. Cochrane Database Syst. Rev. CD001321(2008).
284. Maki, K. C. et al. Consumption of a cranberry juicebeverage lowered the number of clinical urinary tract infection episodes inwomen with a recent history of urinary tract infection. Am. J. Clin. Nutr. 103,1434–1442 (2016).
285. Blumberg, J. B. et al. Cranberries and their bioactiveconstituents in human health. Adv. Nutr. 4, 618–632 (2013).
286. Blumberg, J. B. et al. Impact of Cranberries on GutMicrobiota and Cardiometabolic Health: Proceedings of the Cranberry HealthResearch Conference 2015. Adv. Nutr. 7, 759S–70S (2016).
287. Neto, C. C. & Vinson, J. A. Cranberry. in HerbalMedicine: Biomolecular and Clinical Aspects (eds. Benzie, I. F. F. &Wachtel-Galor, S.) (CRC Press/Taylor & Francis, 2012).
288. Zhang, L. et al. Efficacy of cranberry juice onHelicobacter pylori infection: a double-blind, randomized placebo-controlledtrial. Helicobacter 10, 139–145 (2005).
289. Gotteland, M. et al. Modulation of Helicobacter pyloricolonization with cranberry juice and Lactobacillus johnsonii La1 in children.Nutrition 24, 421–426 (2008).
290. Ruel, G. et al. Favourable impact of low-caloriecranberry juice consumption on plasma HDL-cholesterol concentrations in men.Br. J. Nutr. 96, 357–364 (2006).
291. Lee, I. T., Chan, Y. C., Lin, C. W., Lee, W. J. &Sheu, W. H.-H. Effect of cranberry extracts on lipid profiles in subjects withType 2 diabetes. Diabet. Med. 25, 1473–1477 (2008).
292. Ruel, G. et al. Low-calorie cranberry juicesupplementation reduces plasma oxidized LDL and cell adhesion moleculeconcentrations in men. Br. J. Nutr. 99, 352–359 (2008).
293. Basu, A. et al. Low-energy cranberry juice decreaseslipid oxidation and increases plasma antioxidant capacity in women withmetabolic syndrome. Nutr. Res. 31, 190–196 (2011).
294. Simão, T. N. C. et al. Reduced-energy cranberry juiceincreases folic acid and adiponectin and reduces homocysteine and oxidativestress in patients with the metabolic syndrome. Br. J. Nutr. 110, 1885–1894(2013).
295. Dohadwala, M. M. et al. Effects of cranberry juiceconsumption on vascular function in patients with coronary artery disease. Am.J. Clin. Nutr. 93, 934–940 (2011).
296. Denis, M.-C. et al. Prevention of oxidative stress,inflammation and mitochondrial dysfunction in the intestine by differentcranberry phenolic fractions. Clin. Sci. 128, 197–212 (2015).
297. Caldas, A. P. S., Coelho, O. G. L. & Bressan, J.Cranberry antioxidant power on oxidative stress, inflammation and mitochondrialdamage. Int. J. Food Prop. 21, 582–592 (2018).